

The effect of context on the folding of β-hairpins
- PMID: 21843644
- PMCID: PMC3197233
- DOI: 10.1016/j.jsb.2011.08.001
The effect of context on the folding of β-hairpins
Abstract
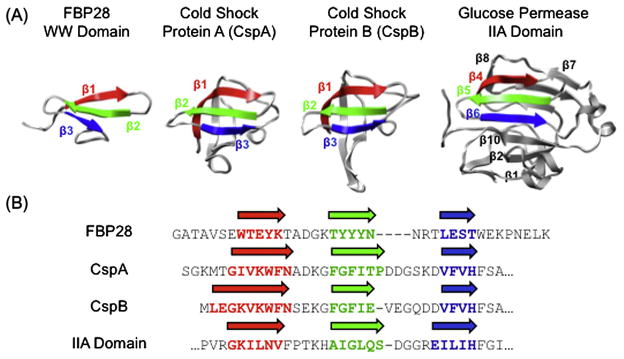
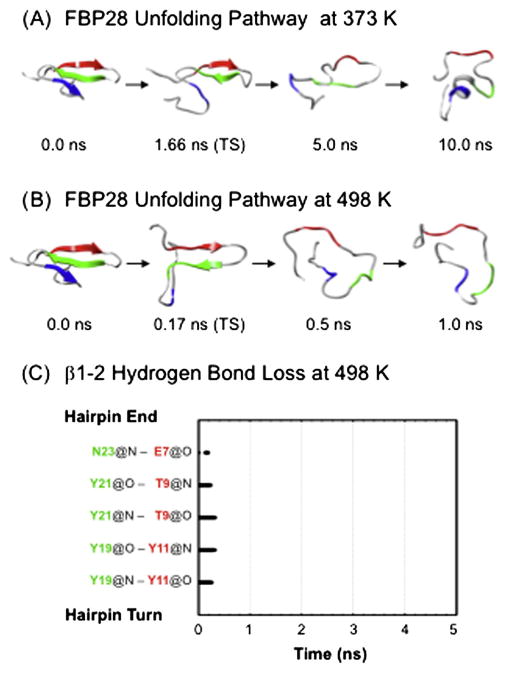
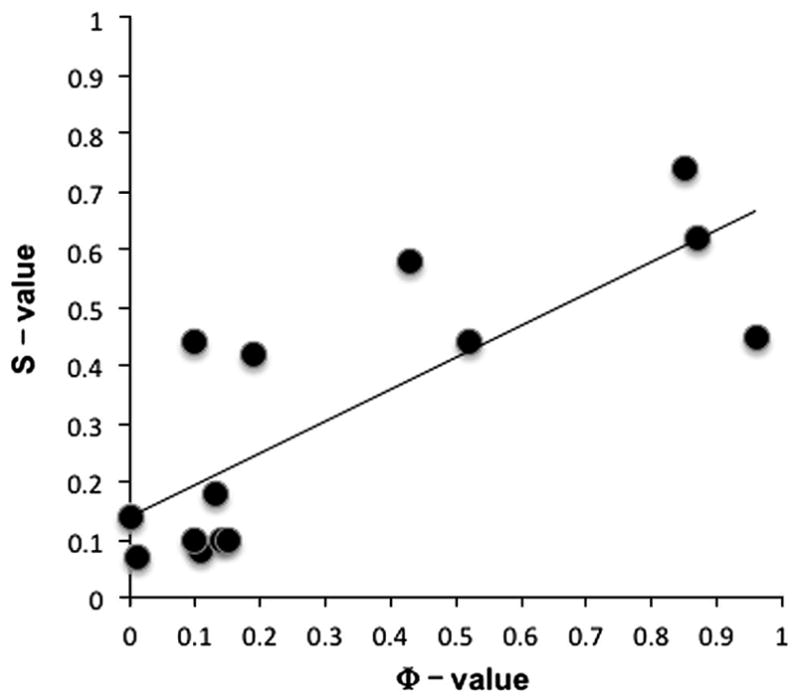
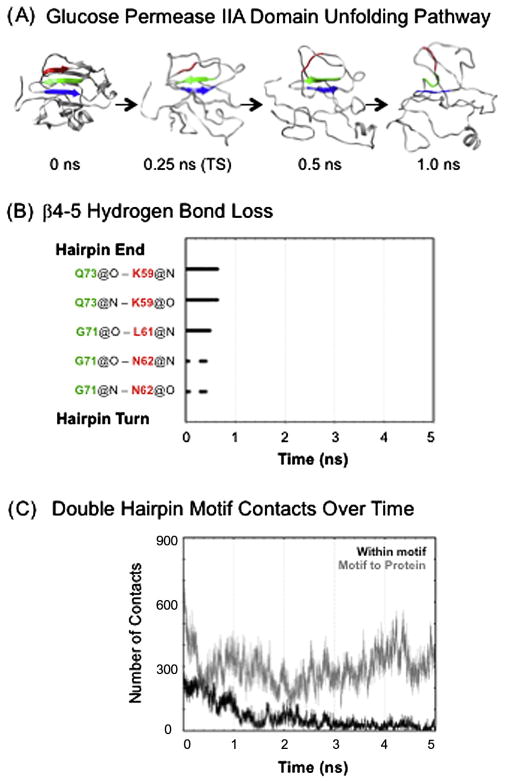
Small β-hairpin peptides have been widely used as models for the folding of β-sheets. But how applicable is the folding of such models to β-structure in larger proteins with conventional hydrophobic cores? Here we present multiple unfolding simulations of three such proteins that contain the WW domain double hairpin β-sheet motif: cold shock protein A (CspA), cold shock protein B (CspB) and glucose permease IIA domain. We compare the behavior of the free motif in solution and in the context of proteins of different size and architecture. Both Csp proteins lost contacts between the double-hairpin motif and the protein core as the first step of unfolding and proceeded to unfold with loss of the third β-strand, similar to the isolated WW domain. The glucose permease IIA domain is a larger protein and the contacts between the motif and the core were not lost as quickly. Instead the unfolding pathway of glucose permease IIA followed a different pathway with β1 pulling away from the sheet first. Interestingly, when the double hairpin motif was excised from the glucose permease IIA domain and simulated in isolation in water it unfolded by the same pathway as the WW domain, indicating that it is tertiary interactions with the protein that alter the motif's unfolding not a sequence dependent effect on its intrinsic unfolding behavior. With respect to the unfolding of the hairpins, there was no consistent order to the loss of hydrogen bonds between the β-strands in the hairpins in any of the systems. Our results show that while the folding behavior of the isolated WW domain is generally consistent with the double hairpin motif's behavior in the cold shock proteins, it is not the case for the glucose permease IIA domain. So, one must be cautious in extrapolating findings from model systems to larger more complicated proteins where tertiary interactions can overwhelm intrinsic behavior.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures



References
-
- Beck DAC, Alonso DOV, Daggett V. in lucem Molecular Mechanics (ilmm) University of Washington; Seattle: 2000–2011.
-
- Beck DAC, Daggett V. Methods for molecular dynamics simulations of protein folding/unfolding in solution. Methods Enzymol. 2004;34:112–120. - PubMed
-
- Beck DAC, Armen RS, Daggett V. Cutoff size need not strongly influence molecular dynamics results for solvated polypeptides. Biochemistry. 2005;44:609–616. - PubMed
-
- Beck DAC, Jonsson AL, Schaeffer RD, Scott KA, Day R, et al. Dynameomics: Mass annotation of protein dynamics and unfolding in water by high-throughput atomistic molecular dynamics simulations. Protein Eng, Des Sel. 2008;21:353–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
