

An iron homeostasis regulatory circuit with reciprocal roles in Candida albicans commensalism and pathogenesis
- PMID: 21843869
- PMCID: PMC3165008
- DOI: 10.1016/j.chom.2011.07.005
An iron homeostasis regulatory circuit with reciprocal roles in Candida albicans commensalism and pathogenesis
Abstract
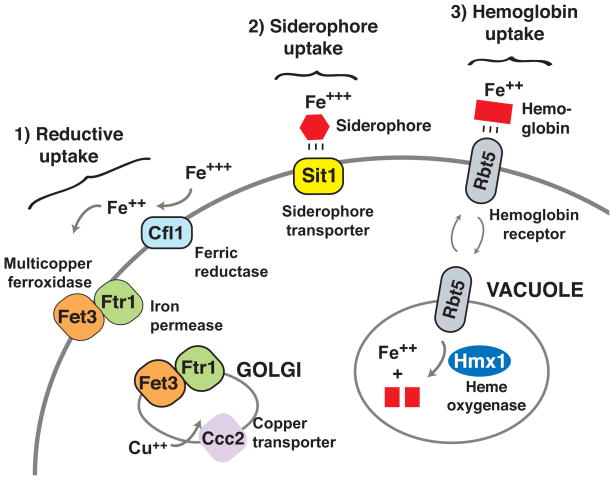
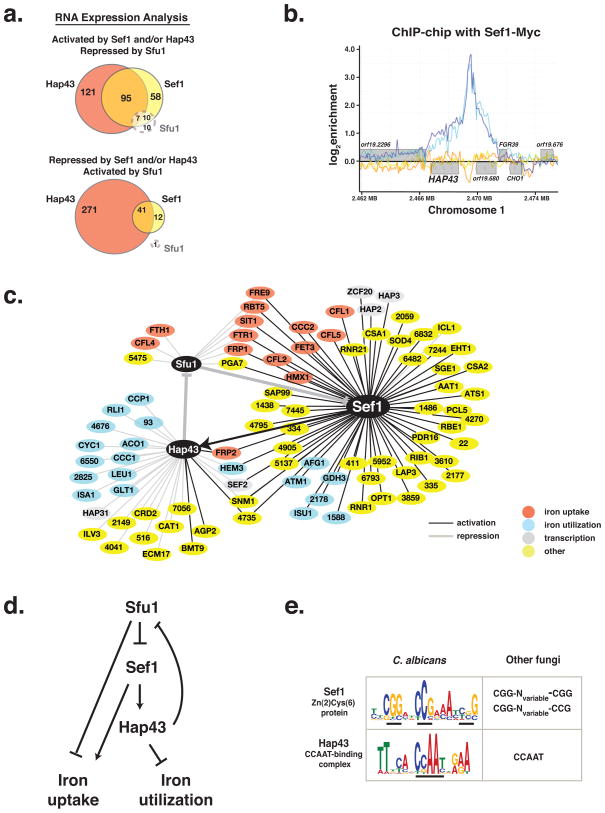
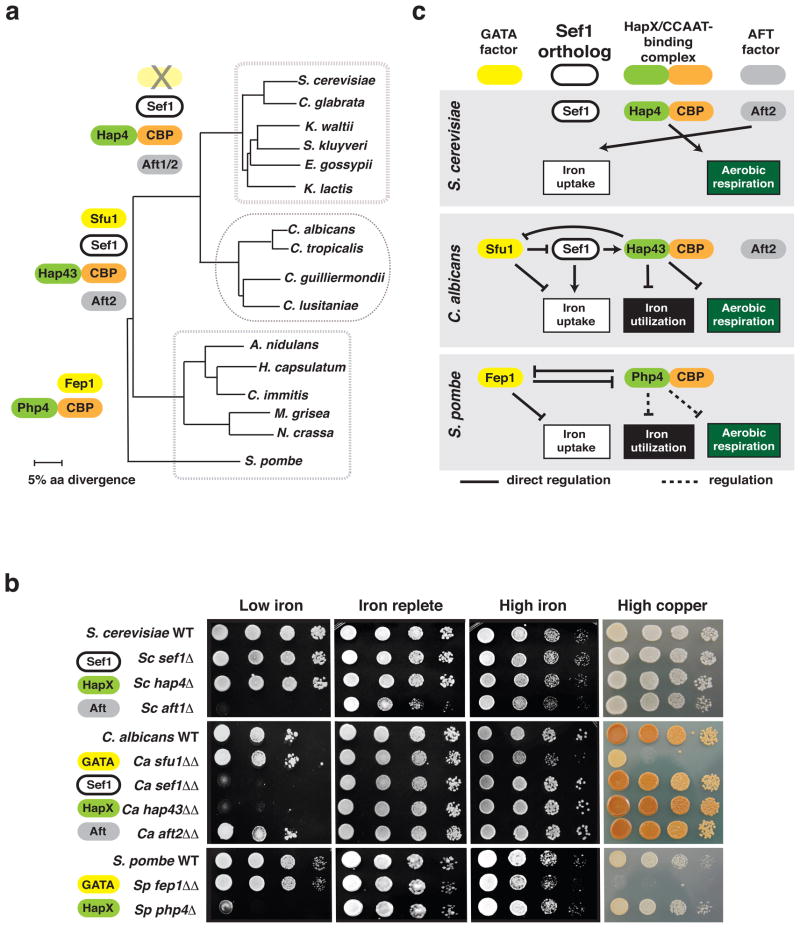
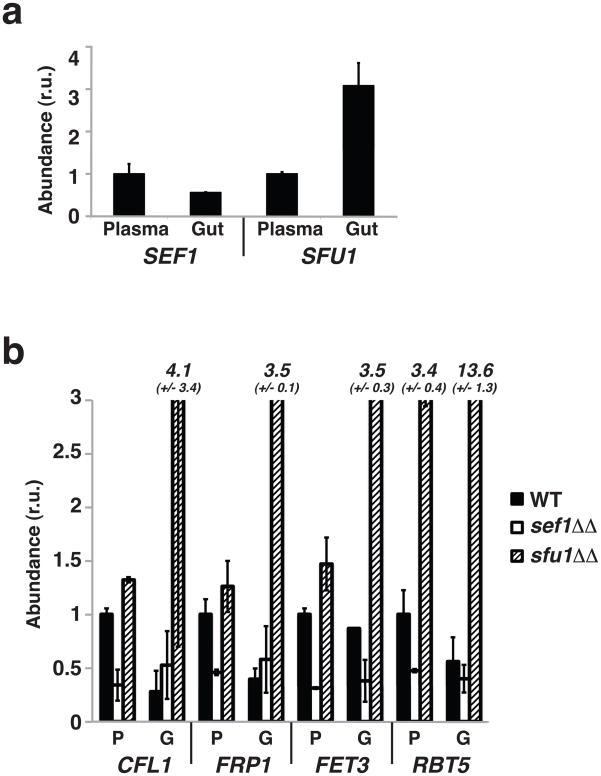
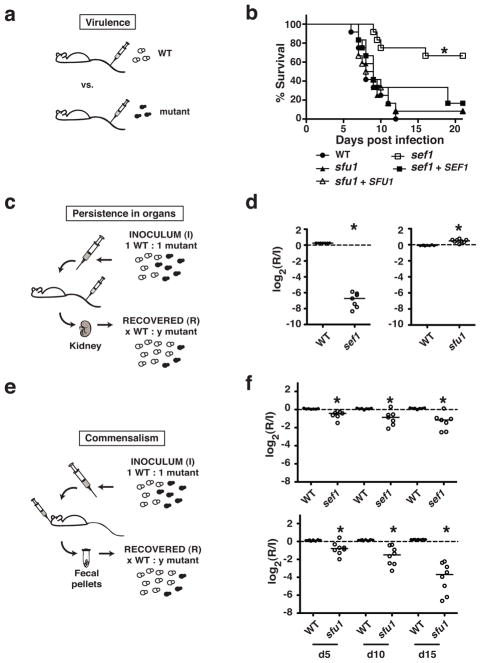
The mammalian gastrointestinal tract and bloodstream are highly disparate biological niches that differ in concentrations of nutrients such as iron. However, some commensal-pathogenic microorganisms, such as the yeast Candida albicans, thrive in both environments. We report the evolution of a transcription circuit in C. albicans that controls iron uptake and determines its fitness in both niches. Our analysis of DNA-binding proteins that regulate iron uptake by this organism suggests the evolutionary intercalation of a transcriptional activator called Sef1 between two broadly conserved iron-responsive transcriptional repressors, Sfu1 and Hap43. Sef1 activates iron-uptake genes and promotes virulence in a mouse model of bloodstream infection, whereas Sfu1 represses iron-uptake genes and is dispensable for virulence but promotes gastrointestinal commensalism. Thus, C. albicans can alternate between genetic programs conferring resistance to iron depletion in the bloodstream versus iron toxicity in the gut, and this may represent a fundamental attribute of gastrointestinal commensal-pathogens.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Candida albicans adds more weight to iron regulation.Cell Host Microbe. 2011 Aug 18;10(2):93-4. doi: 10.1016/j.chom.2011.08.001. Cell Host Microbe. 2011. PMID: 21843865
References
-
- Ardon O, Bussey H, Philpott C, Ward DM, Davis-Kaplan S, Verroneau S, Jiang B, Kaplan J. Identification of a Candida albicans ferrichrome transporter and its characterization by expression in Saccharomyces cerevisiae. J Biol Chem. 2001;276:43049–43055. - PubMed
-
- Bailey TL, Elkan C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Paper presented at: Second International Conference on Intelligent Systems for Molecular Biology (AAAI Press).1994. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
