

The Paf oncogene is essential for hematopoietic stem cell function and development
- PMID: 21844206
- PMCID: PMC3171089
- DOI: 10.1084/jem.20102170
The Paf oncogene is essential for hematopoietic stem cell function and development
Abstract
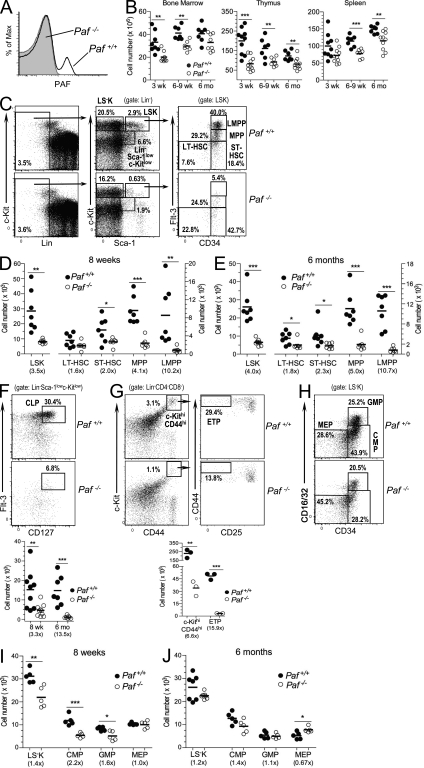
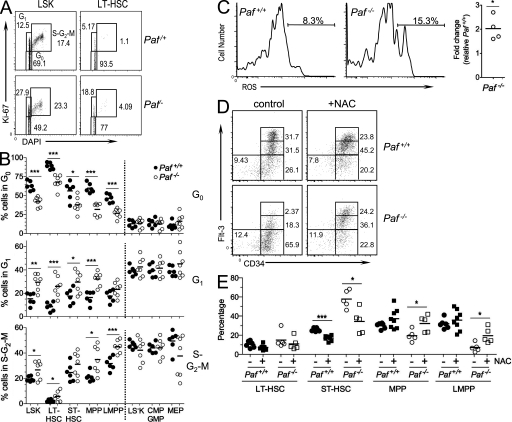
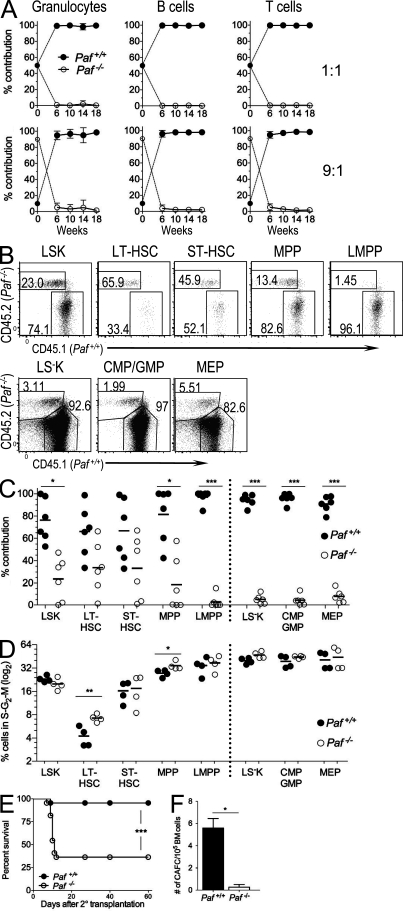
Hematopoietic stem cells (HSCs) self-renew to maintain the lifelong production of all blood populations. Here, we show that the proliferating cell nuclear antigen-associated factor (Paf) is highly expressed in cycling bone marrow HSCs and plays a critical role in hematopoiesis. Mice lacking Paf exhibited reduced bone marrow cellularity; reduced numbers of HSCs and committed progenitors; and leukopenia. These phenotypes are caused by a cell-intrinsic blockage in the development of long-term (LT)-HSCs into multipotent progenitors and preferential loss of lymphoid progenitors caused by markedly increased p53-mediated apoptosis. In addition, LT-HSCs from Paf(-/-) mice had increased levels of reactive oxygen species (ROS), failed to maintain quiescence, and were unable to support LT hematopoiesis. The loss of lymphoid progenitors was likely due the increased levels of ROS in LT-HSCs caused by treatment of Paf(-/-) mice with the anti-oxidant N-acetylcysteine restored lymphoid progenitor numbers to that of Paf(+/+) mice. Collectively, our studies identify Paf as a novel and essential regulator of early hematopoiesis.
© 2011 Amrani et al.
Figures

References
-
- Adolfsson J., Borge O.J., Bryder D., Theilgaard-Mönch K., Astrand-Grundström I., Sitnicka E., Sasaki Y., Jacobsen S.E. 2001. Upregulation of Flt3 expression within the bone marrow Lin(-)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity. 15:659–669 10.1016/S1074-7613(01)00220-5 - DOI - PubMed
-
- Breems D.A., Blokland E.A., Neben S., Ploemacher R.E. 1994. Frequency analysis of human primitive haematopoietic stem cell subsets using a cobblestone area forming cell assay. Leukemia. 8:1095–1104 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
