

OCT4 establishes and maintains nucleosome-depleted regions that provide additional layers of epigenetic regulation of its target genes
- PMID: 21844352
- PMCID: PMC3167525
- DOI: 10.1073/pnas.1111309108
OCT4 establishes and maintains nucleosome-depleted regions that provide additional layers of epigenetic regulation of its target genes
Abstract
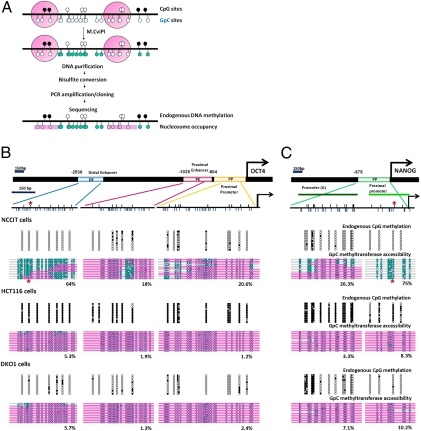
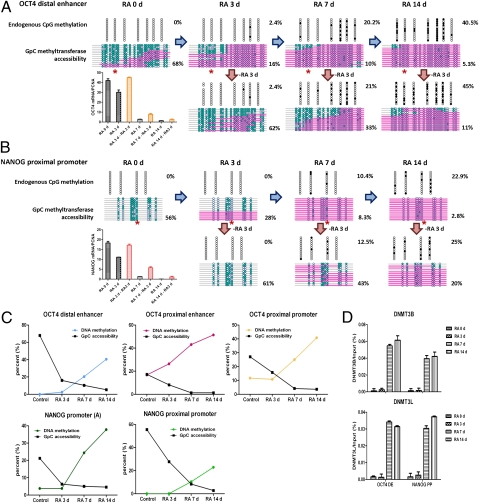
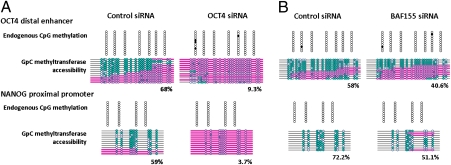
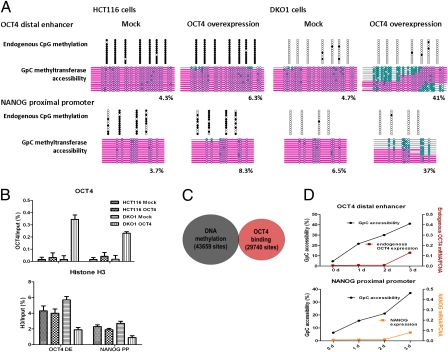
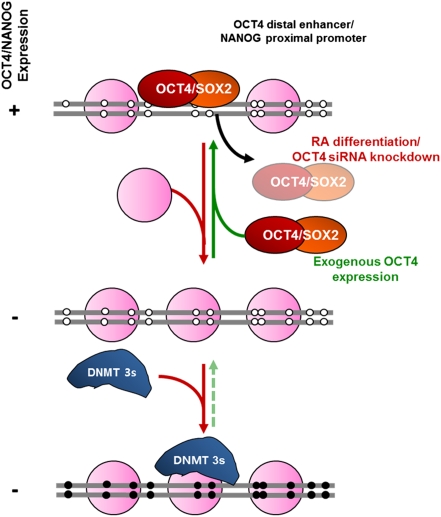
Recent epigenome-wide mapping studies describe nucleosome-depleted regions (NDRs) at transcription start sites and enhancers. However, these static maps do not address causality or the roles of NDRs in gene control, and their relationship to transcription factors and DNA methylation is not well understood. Using a high-resolution single-molecule mapping approach to simultaneously investigate endogenous DNA methylation and nucleosome occupancies on individual DNA molecules, we show that the unmethylated OCT4 distal enhancer has an NDR, whereas NANOG has a clear NDR at its proximal promoter. These NDRs are maintained by binding of OCT4 and are required for OCT4 and NANOG expression. Differentiation causes a rapid loss of both NDRs accompanied by nucleosome occupancy, which precedes de novo DNA methylation. NDRs can be restored by forced expression of OCT4 in somatic cells but only when there is no cytosine methylation. These data show the central role of the NDRs, established by OCT4, in ensuring the autoregulatory loop of pluripotency and, furthermore, that de novo methylation follows the loss of NDRs and stabilizes the suppressed state.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
