

Defining the nature of human pluripotent stem cell progeny
- PMID: 21844894
- PMCID: PMC3351932
- DOI: 10.1038/cr.2011.133
Defining the nature of human pluripotent stem cell progeny
Abstract
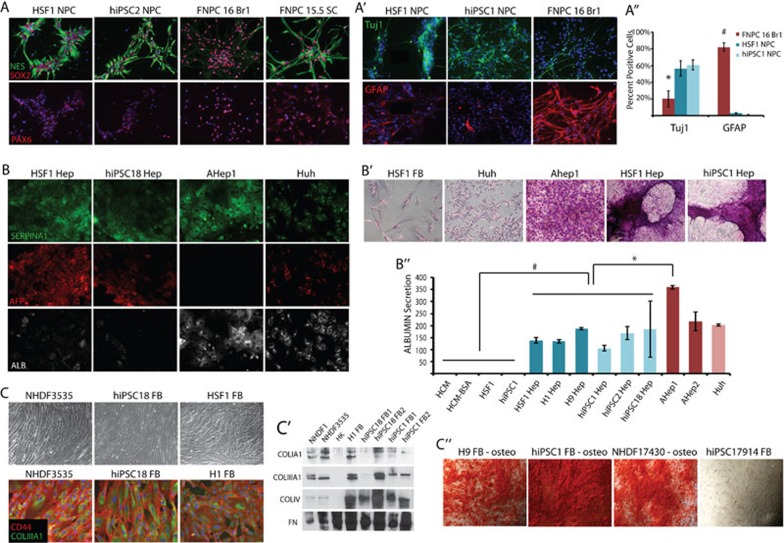
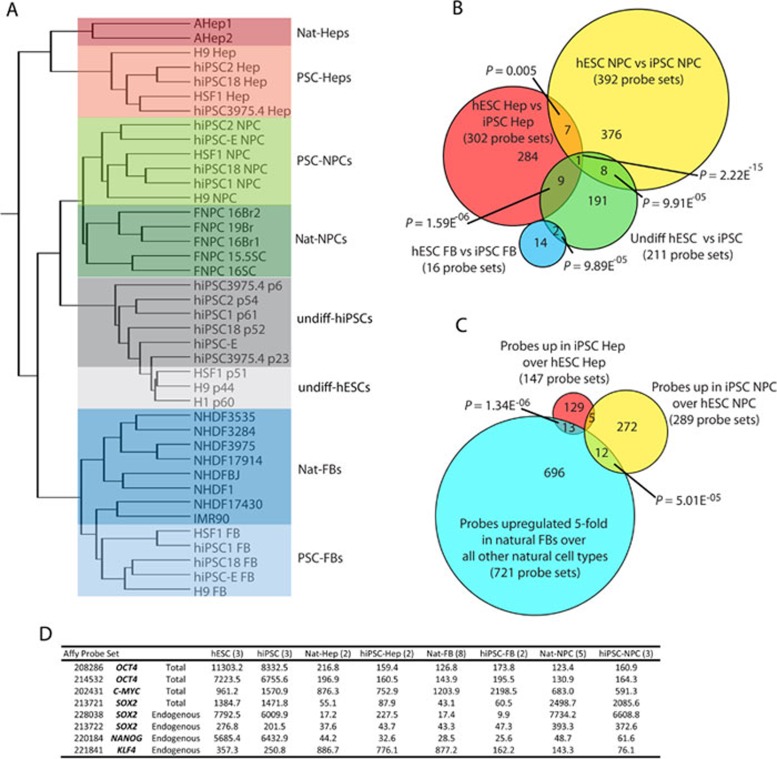
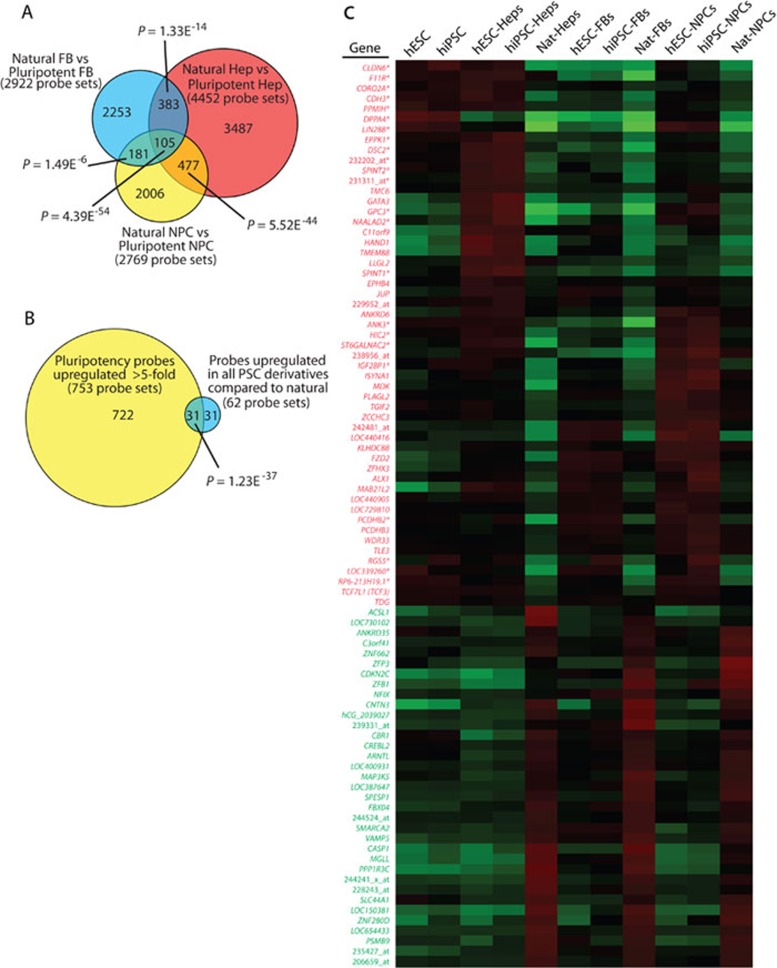
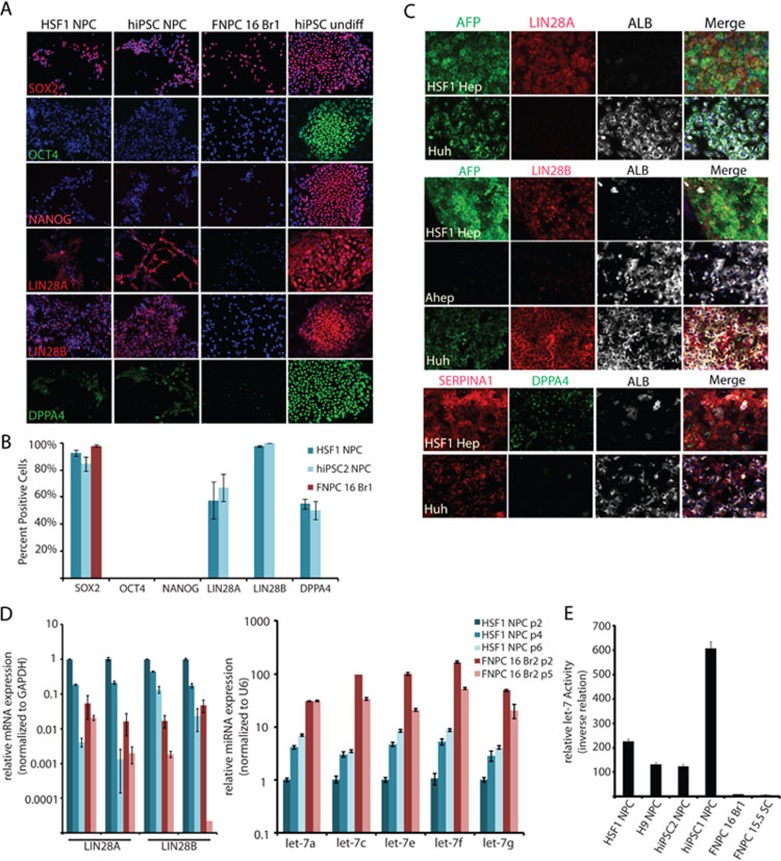
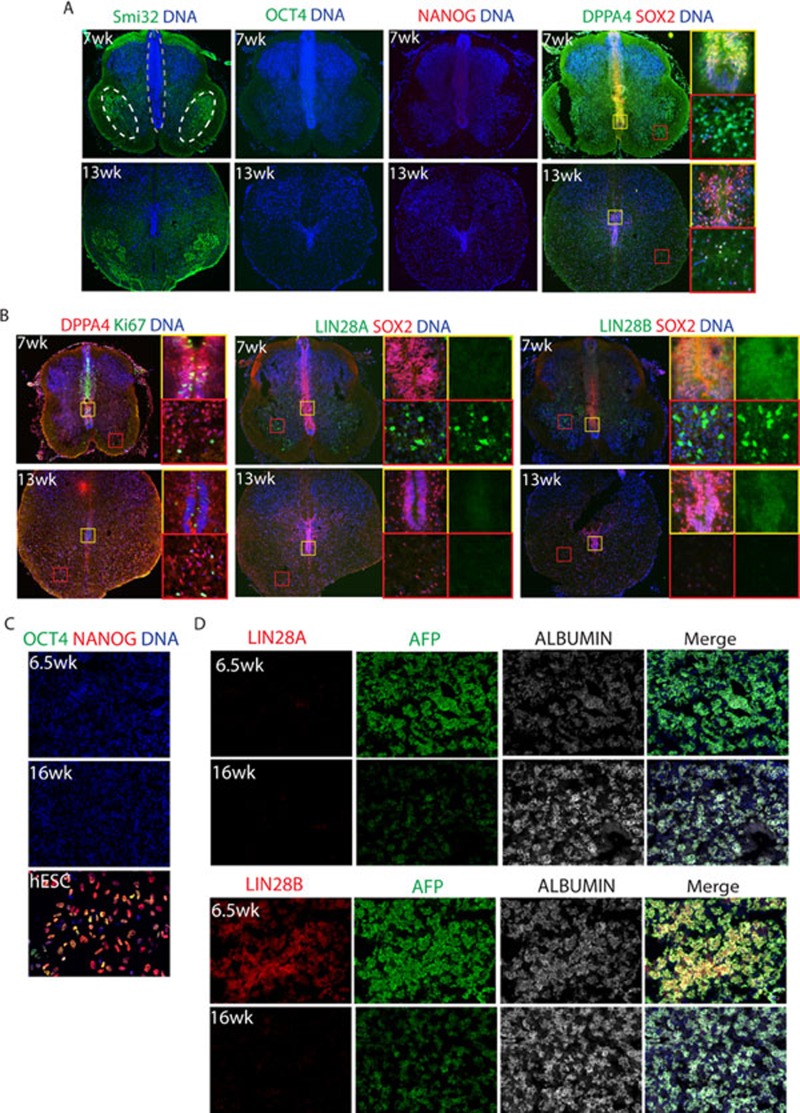
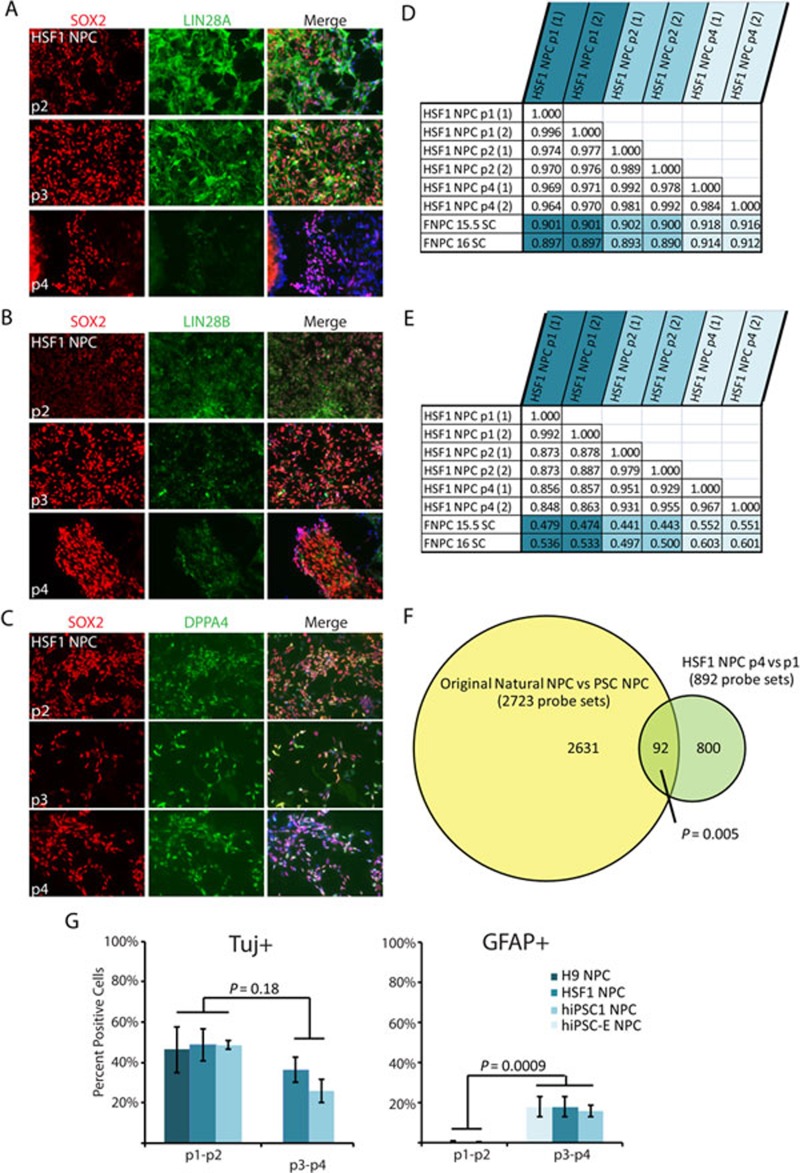
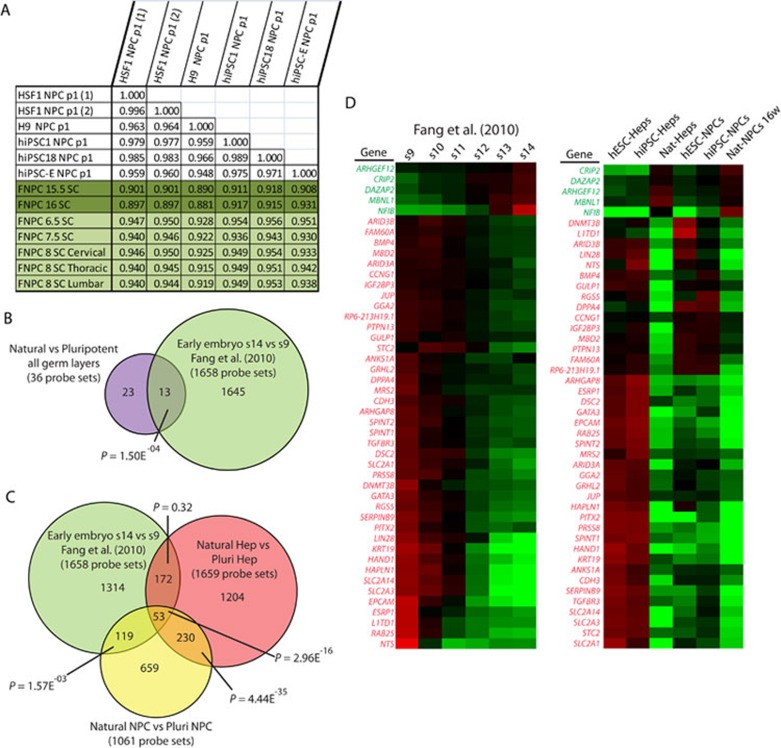
While it is clear that human pluripotent stem cells (hPSCs) can differentiate to generate a panoply of various cell types, it is unknown how closely in vitro development mirrors that which occurs in vivo. To determine whether human embryonic stem cells (hESCs) and human-induced pluripotent stem cells (hiPSCs) make equivalent progeny, and whether either makes cells that are analogous to tissue-derived cells, we performed comprehensive transcriptome profiling of purified PSC derivatives and their tissue-derived counterparts. Expression profiling demonstrated that hESCs and hiPSCs make nearly identical progeny for the neural, hepatic, and mesenchymal lineages, and an absence of re-expression from exogenous reprogramming factors in hiPSC progeny. However, when compared to a tissue-derived counterpart, the progeny of both hESCs and hiPSCs maintained expression of a subset of genes normally associated with early mammalian development, regardless of the type of cell generated. While pluripotent genes (OCT4, SOX2, REX1, and NANOG) appeared to be silenced immediately upon differentiation from hPSCs, genes normally unique to early embryos (LIN28A, LIN28B, DPPA4, and others) were not fully silenced in hPSC derivatives. These data and evidence from expression patterns in early human fetal tissue (3-16 weeks of development) suggest that the differentiated progeny of hPSCs are reflective of very early human development (< 6 weeks). These findings provide support for the idea that hPSCs can serve as useful in vitro models of early human development, but also raise important issues for disease modeling and the clinical application of hPSC derivatives.
Figures
References
-
- Takahashi K, Tanabe K, Ohnuki M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
