

Mechanism of AMPA receptor activation by partial agonists: disulfide trapping of closed lobe conformations
- PMID: 21846932
- PMCID: PMC3186401
- DOI: 10.1074/jbc.M111.269001
Mechanism of AMPA receptor activation by partial agonists: disulfide trapping of closed lobe conformations
Abstract
The mechanism by which agonist binding to an ionotropic glutamate receptor leads to channel opening is a central issue in molecular neurobiology. Partial agonists are useful tools for studying the activation mechanism because they produce full channel activation with lower probability than full agonists. Structural transitions that determine the efficacy of partial agonists can provide information on the trigger that begins the channel-opening process. The ligand-binding domain of AMPA receptors is a bilobed structure, and the closure of the lobes is associated with channel activation. One possibility is that partial agonists sterically block full lobe closure but that partial degrees of closure trigger the channel with a lower probability. Alternatively, full lobe closure may be required for activation, and the stability of the fully closed state could determine efficacy with the fully closed state having a lower stability when bound to partial relative to full agonists. Disulfide-trapping experiments demonstrated that even extremely low efficacy ligands such as 6-cyano-7-nitroquinoxaline-2,3-dione can produce a full lobe closure, presumably with low probability. The results are consistent the hypothesis that the efficacy is determined at least in part by the stability of the state in which the lobes are fully closed.
Figures
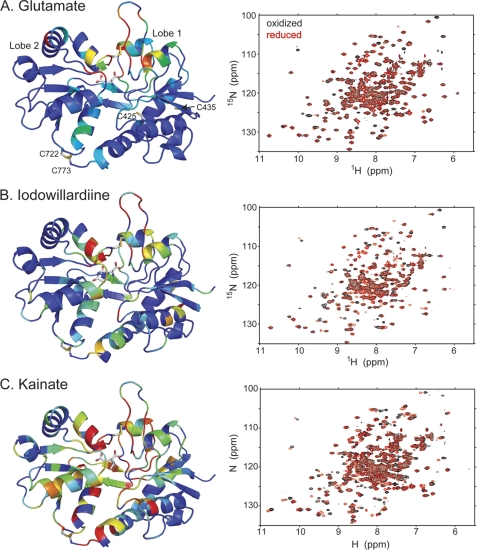
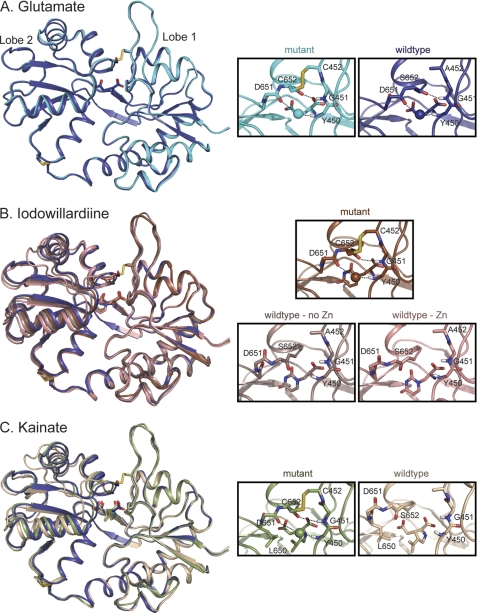
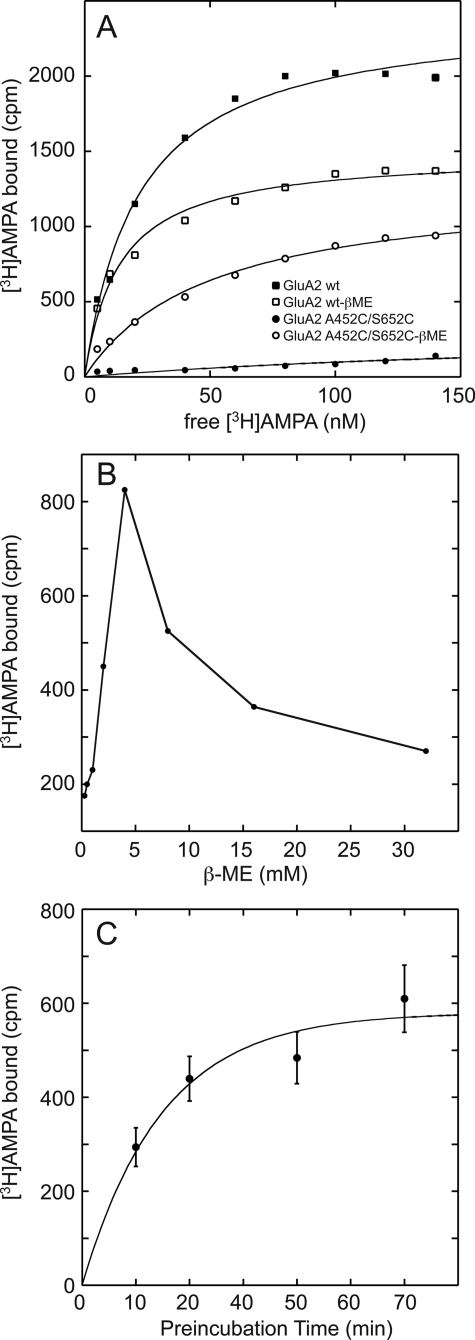
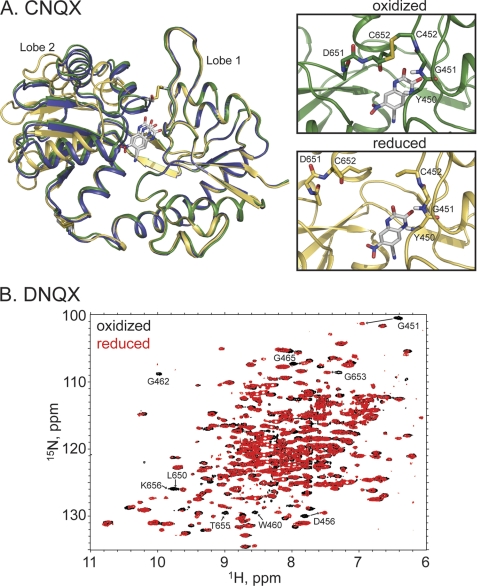
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
