

CBP is required for environmental enrichment-induced neurogenesis and cognitive enhancement
- PMID: 21847097
- PMCID: PMC3199387
- DOI: 10.1038/emboj.2011.299
CBP is required for environmental enrichment-induced neurogenesis and cognitive enhancement
Abstract
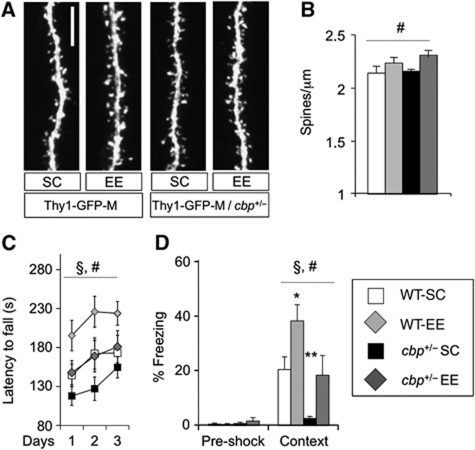
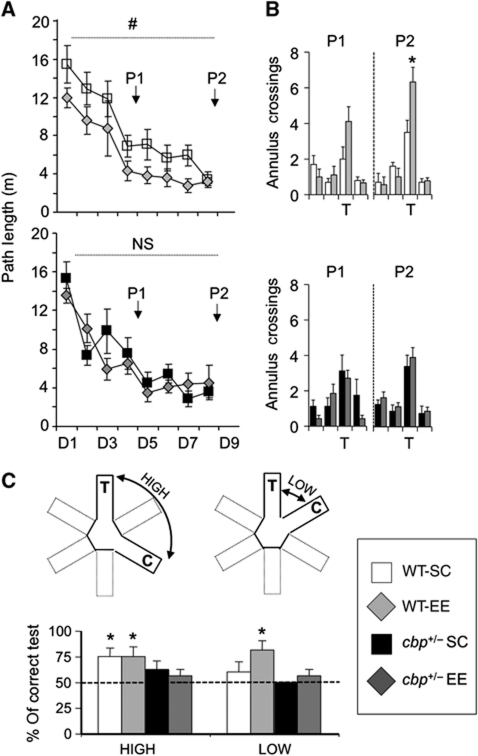
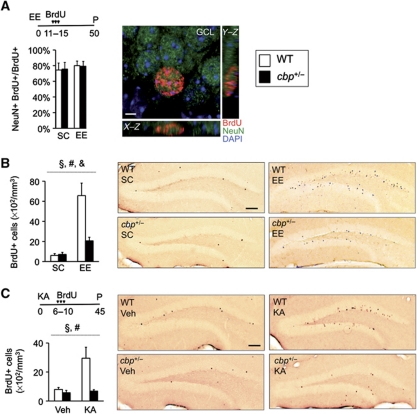
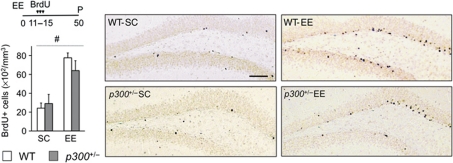
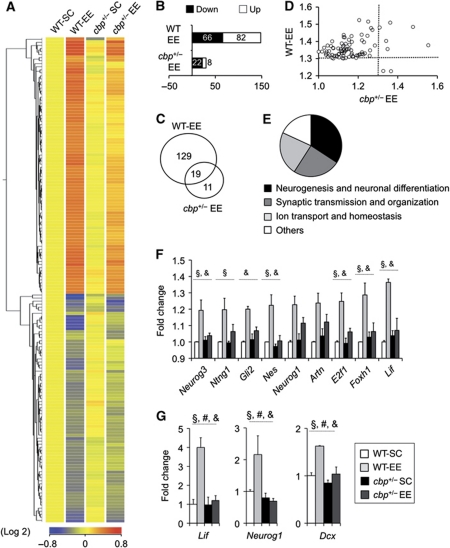
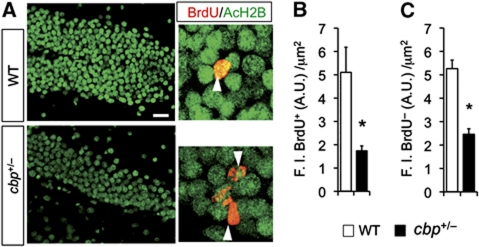
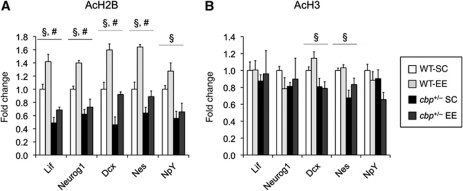
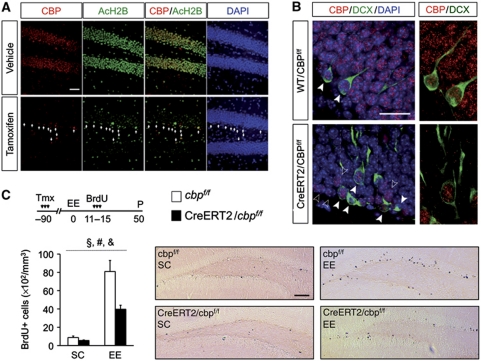
The epigenetic changes of the chromatin represent an attractive molecular substrate for adaptation to the environment. We examined here the role of CREB-binding protein (CBP), a histone acetyltransferase involved in mental retardation, in the genesis and maintenance of long-lasting systemic and behavioural adaptations to environmental enrichment (EE). Morphological and behavioural analyses demonstrated that EE ameliorates deficits associated to CBP deficiency. However, CBP-deficient mice also showed a strong defect in environment-induced neurogenesis and impaired EE-mediated enhancement of spatial navigation and pattern separation ability. These defects correlated with an attenuation of the transcriptional programme induced in response to EE and with deficits in histone acetylation at the promoters of EE-regulated, neurogenesis-related genes. Additional experiments in CBP restricted and inducible knockout mice indicated that environment-induced adult neurogenesis is extrinsically regulated by CBP function in mature granule cells. Overall, our experiments demonstrate that the environment alters gene expression by impinging on activities involved in modifying the epigenome and identify CBP-dependent transcriptional neuroadaptation as an important mediator of EE-induced benefits, a finding with important implications for mental retardation therapeutics.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Alarcon JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A (2004) Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 42: 947–959 - PubMed
-
- Barco A (2007) The Rubinstein-Taybi syndrome: modeling mental impairment in the mouse. Genes Brain Behav 6: 32–39 - PubMed
