

Clathrin light chain directs endocytosis by influencing the binding of the yeast Hip1R homologue, Sla2, to F-actin
- PMID: 21849475
- PMCID: PMC3183023
- DOI: 10.1091/mbc.E11-07-0628
Clathrin light chain directs endocytosis by influencing the binding of the yeast Hip1R homologue, Sla2, to F-actin
Abstract
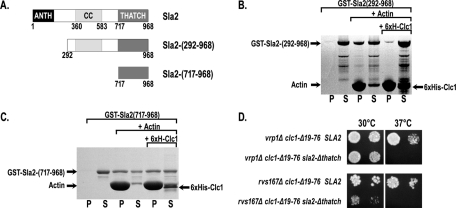
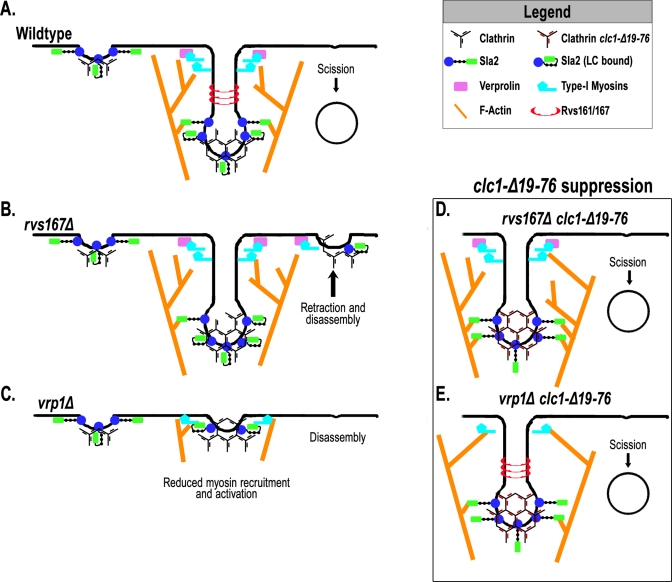
The role of clathrin light chain (CLC) in clathrin-mediated endocytosis is not completely understood. Previous studies showed that the CLC N-terminus (CLC-NT) binds the Hip1/Hip1R/Sla2 family of membrane/actin-binding factors and that overexpression of the CLC-NT in yeast suppresses endocytic defects of clathrin heavy-chain mutants. To elucidate the mechanistic basis for this suppression, we performed synthetic genetic array analysis with a clathrin CLC-NT deletion mutation (clc1-Δ19-76). clc1-Δ19-76 suppressed the internalization defects of null mutations in three late endocytic factors: amphiphysins (rvs161 and rvs167) and verprolin (vrp1). In actin sedimentation assays, CLC binding to Sla2 inhibited Sla2 interaction with F-actin. Furthermore, clc1-Δ19-76 suppression of the rvs and vrp phenotypes required the Sla2 actin-binding talin-Hip1/R/Sla2 actin-tethering C-terminal homology domain, suggesting that clc1-Δ19-76 promotes internalization by prolonging actin engagement by Sla2. We propose that CLC directs endocytic progression by pruning the Sla2-actin attachments in the clathrin lattice, providing direction for membrane internalization.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
