

Unique ganglioside recognition strategies for clostridial neurotoxins
- PMID: 21849494
- PMCID: PMC3190786
- DOI: 10.1074/jbc.M111.272054
Unique ganglioside recognition strategies for clostridial neurotoxins
Abstract
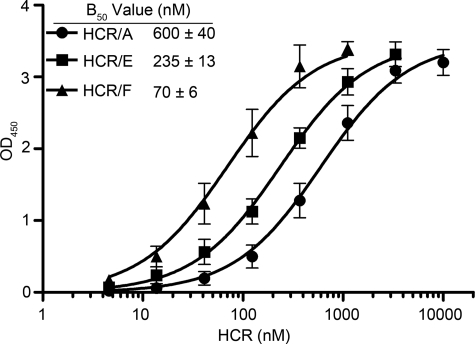
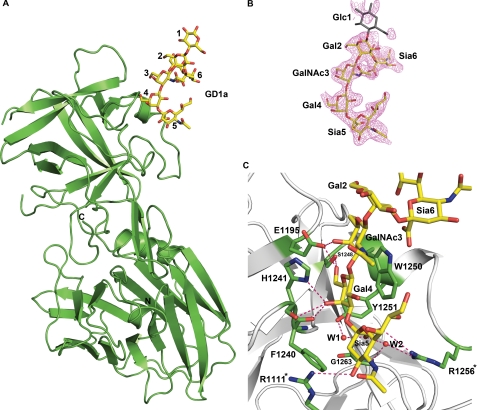
Botulinum neurotoxins (BoNTs) and tetanus neurotoxin are the causative agents of the paralytic diseases botulism and tetanus, respectively. The potency of the clostridial neurotoxins (CNTs) relies primarily on their highly specific binding to nerve terminals and cleavage of SNARE proteins. Although individual CNTs utilize distinct proteins for entry, they share common ganglioside co-receptors. Here, we report the crystal structure of the BoNT/F receptor-binding domain in complex with the sugar moiety of ganglioside GD1a. GD1a binds in a shallow groove formed by the conserved peptide motif E … H … SXWY … G, with additional stabilizing interactions provided by two arginine residues. Comparative analysis of BoNT/F with other CNTs revealed several differences in the interactions of each toxin with ganglioside. Notably, exchange of BoNT/F His-1241 with the corresponding lysine residue of BoNT/E resulted in increased affinity for GD1a and conferred the ability to bind ganglioside GM1a. Conversely, BoNT/E was not able to bind GM1a, demonstrating a discrete mechanism of ganglioside recognition. These findings provide a structural basis for ganglioside binding among the CNTs and show that individual toxins utilize unique ganglioside recognition strategies.
Figures

References
-
- Arnon S. S., Schechter R., Inglesby T. V., Henderson D. A., Bartlett J. G., Ascher M. S., Eitzen E., Fine A. D., Hauer J., Layton M., Lillibridge S., Osterholm M. T., O'Toole T., Parker G., Perl T. M., Russell P. K., Swerdlow D. L., Tonat K. (2001) JAMA 285, 1059–1070 - PubMed
-
- Johnson E. A. (1999) Annu. Rev. Microbiol. 53, 551–575 - PubMed
-
- Johnson E. A., Bradshaw M. (2001) Toxicon 39, 1703–1722 - PubMed
-
- Hatheway C. L. (1995) Curr. Top. Microbiol. Immunol. 195, 55–75 - PubMed
-
- Baldwin M. R., Kim J. J., Barbieri J. T. (2007) Nat. Struct. Mol. Bio.l 14, 9–10 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
