

Dynamic interactions between transposable elements and their hosts
- PMID: 21850042
- PMCID: PMC3192332
- DOI: 10.1038/nrg3030
Dynamic interactions between transposable elements and their hosts
Abstract
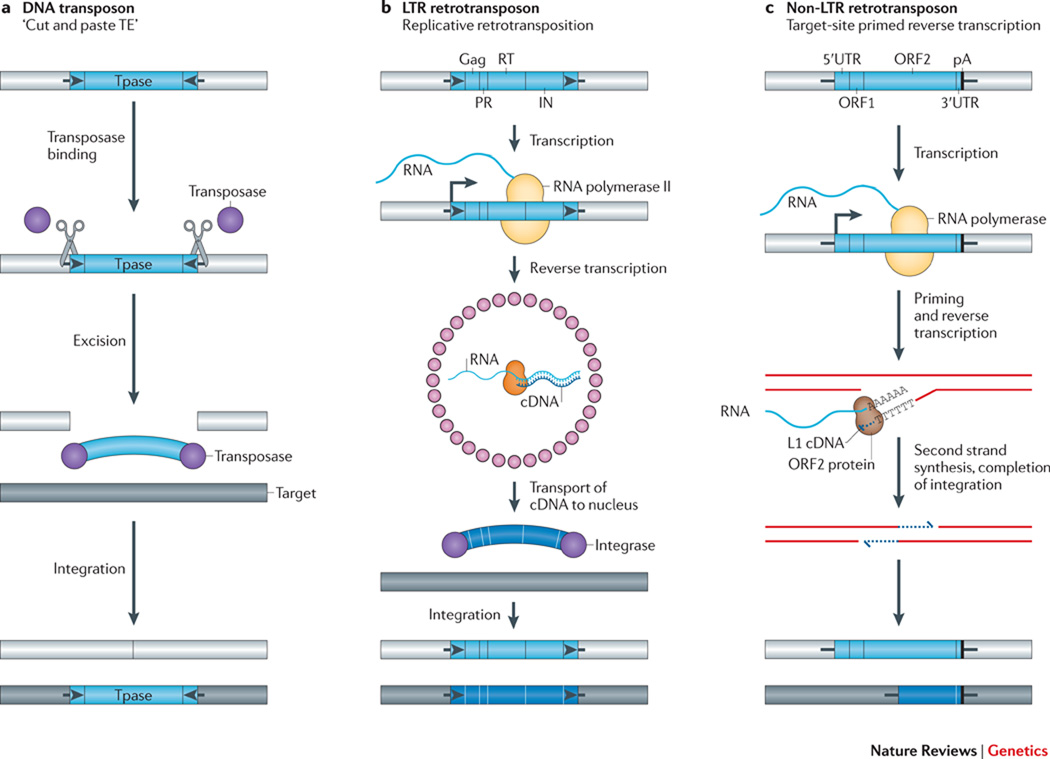
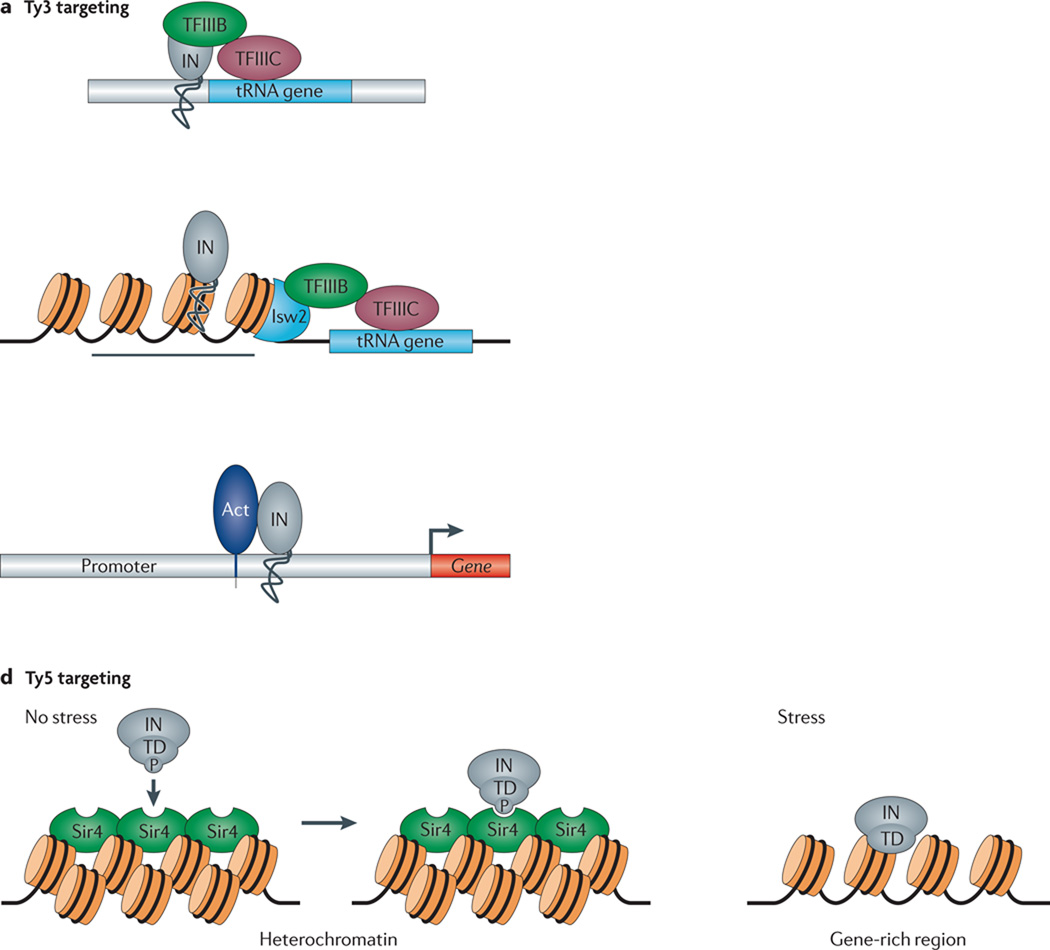
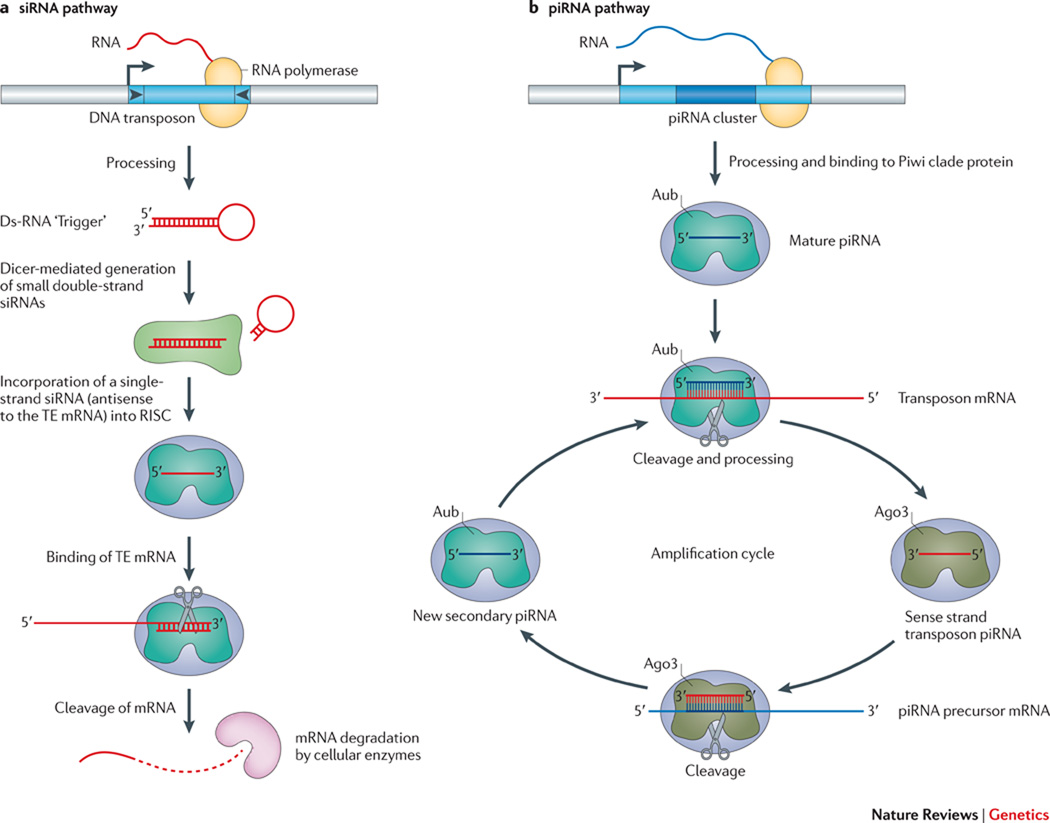
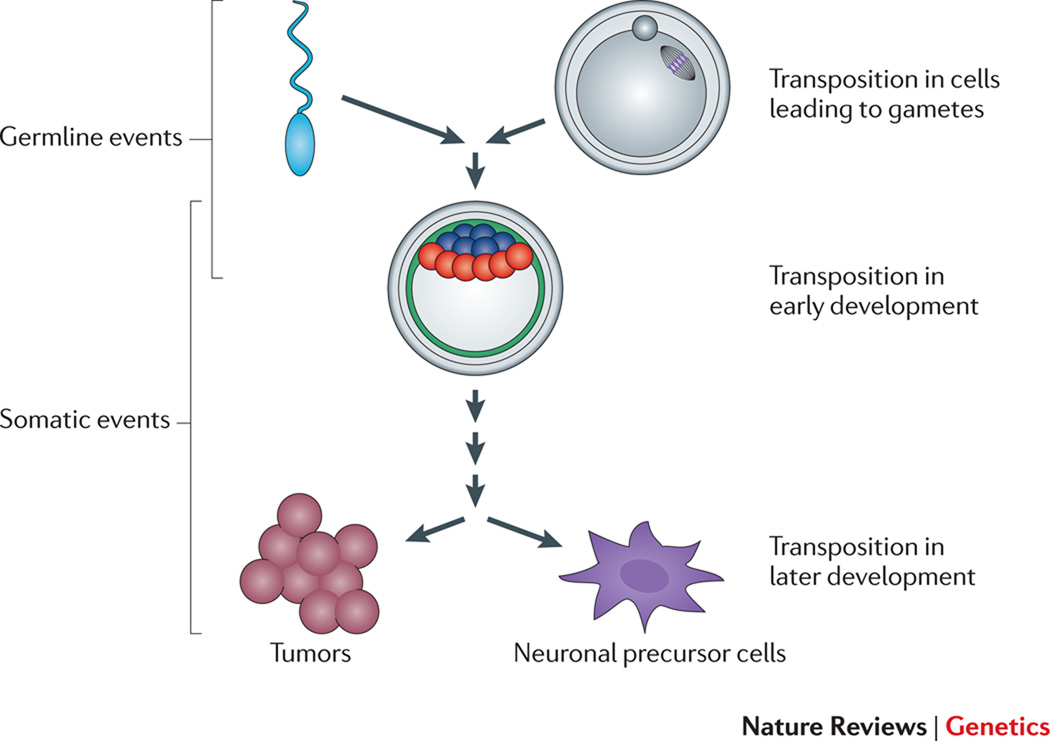
Transposable elements (TEs) have a unique ability to mobilize to new genomic locations, and the major advance of second-generation DNA sequencing has provided insights into the dynamic relationship between TEs and their hosts. It now is clear that TEs have adopted diverse strategies - such as specific integration sites or patterns of activity - to thrive in host environments that are replete with mechanisms, such as small RNAs or epigenetic marks, that combat TE amplification. Emerging evidence suggests that TE mobilization might sometimes benefit host genomes by enhancing genetic diversity, although TEs are also implicated in diseases such as cancer. Here, we discuss recent findings about how, where and when TEs insert in diverse organisms.
Figures
References
-
- Levis RW, Ganesan R, Houtchens K, Tolar LA, Sheen FM. Transposons in place of telomeric repeats at a Drosophila telomere. Cell. 1993;75:1083–1093. - PubMed
-
- Agrawal A, Eastman QM, Schatz DG. Transposition mediated by RAG1 and RAG2 and its implications for the evolution of the immune system. Nature. 1998;394:744–751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
