

Transforming growth factor-β regulates the growth of valve interstitial cells in vitro
- PMID: 21851806
- PMCID: PMC3181399
- DOI: 10.1016/j.ajpath.2011.06.007
Transforming growth factor-β regulates the growth of valve interstitial cells in vitro
Abstract
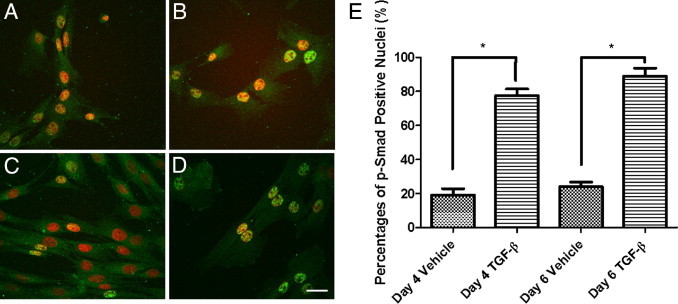
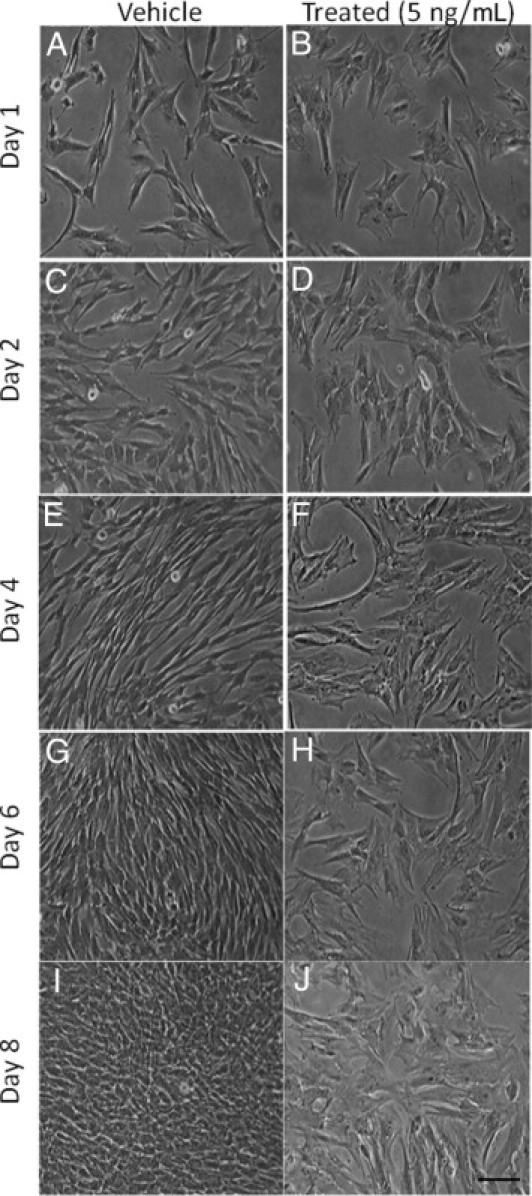
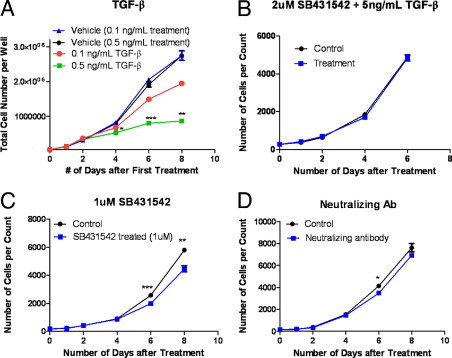
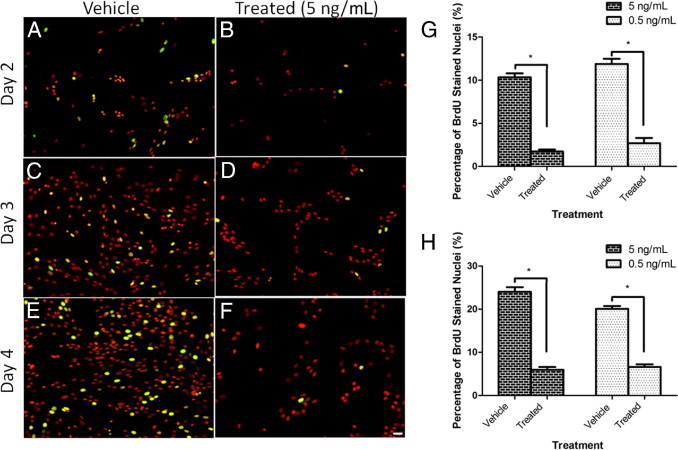
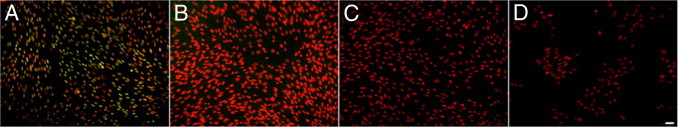
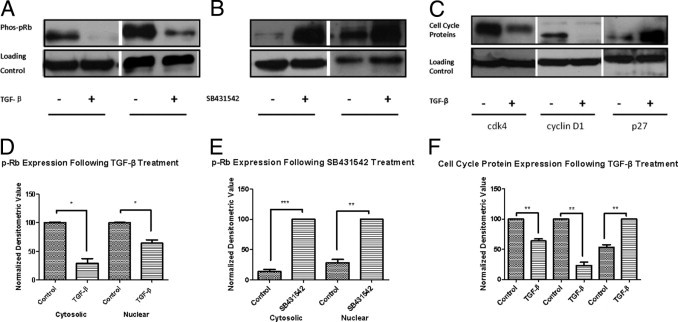
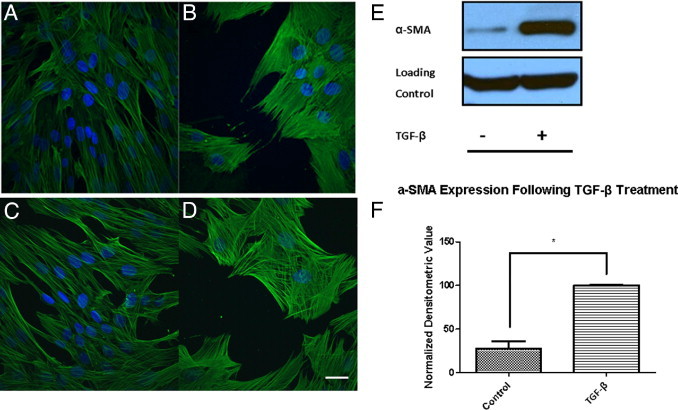
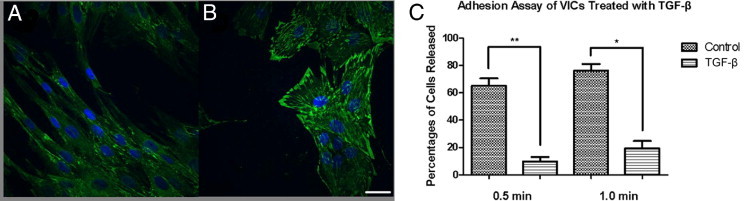
Although valve interstitial cell (VIC) growth is an essential feature of injured and diseased valves, the regulation of VIC growth is poorly understood. Transforming growth factor (TGF)-β promotes VIC proliferation in early-stage wound repair; thus, herein, we tested the hypothesis that TGF-β regulates VIC proliferation under normal nonwound conditions using low-density porcine VIC monolayers. Cell numbers were counted during a 10-day period, whereas proliferation and apoptosis were quantified by bromodeoxyuridine staining and TUNEL, respectively. The extent of retinoblastoma protein phosphorylation and expression of cyclin D1, CDK 4, and p27 were compared using Western blot analysis. Adhesion was quantified using a trypsin adhesion assay, and morphological change was demonstrated by immunofluorescence localization of α-smooth muscle actin and vinculin. TGF-β-treated VICs were rhomboid; significantly decreased in number, proliferation, and retinoblastoma protein phosphorylation; and concomitantly had decreased expression of cyclin D1/CDK4 and increased expression of p27. TGF-β-treated VICs adhered better to substratum and had more vinculin plaques and α-smooth muscle actin stress fibers than did controls. Thus, the regulation of VIC growth by TGF-β is context dependent. TGF-β prevents excessive heart valve growth under normal physiological conditions while it promotes cell proliferation in the early stages of repair, when increased VICs are required.
Copyright © 2011 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Transforming growth factor-beta regulates in vitro heart valve repair by activated valve interstitial cells.Am J Pathol. 2008 Nov;173(5):1275-85. doi: 10.2353/ajpath.2008.080365. Epub 2008 Oct 2. Am J Pathol. 2008. PMID: 18832581 Free PMC article.
-
Transforming growth factor-β1 promotes fibrosis but attenuates calcification of valvular tissue applied as a three-dimensional calcific aortic valve disease model.Am J Physiol Heart Circ Physiol. 2020 Nov 1;319(5):H1123-H1141. doi: 10.1152/ajpheart.00651.2019. Epub 2020 Sep 28. Am J Physiol Heart Circ Physiol. 2020. PMID: 32986963
-
Wnt3a/β-catenin increases proliferation in heart valve interstitial cells.Cardiovasc Pathol. 2013 Mar-Apr;22(2):156-66. doi: 10.1016/j.carpath.2012.06.008. Epub 2012 Aug 11. Cardiovasc Pathol. 2013. PMID: 22889676
-
On the Functional Role of Valve Interstitial Cell Stress Fibers: A Continuum Modeling Approach.J Biomech Eng. 2017 Feb 1;139(2):0210071-02100713. doi: 10.1115/1.4035557. J Biomech Eng. 2017. PMID: 28024085 Free PMC article. Review.
-
Cyclin-dependent kinase regulation during G1 phase and cell cycle regulation by TGF-beta.Adv Cancer Res. 1997;71:165-207. doi: 10.1016/s0065-230x(08)60099-8. Adv Cancer Res. 1997. PMID: 9111866 Review.
Cited by
-
Mitral valve disease--morphology and mechanisms.Nat Rev Cardiol. 2015 Dec;12(12):689-710. doi: 10.1038/nrcardio.2015.161. Epub 2015 Oct 20. Nat Rev Cardiol. 2015. PMID: 26483167 Free PMC article. Review.
-
TGF-β signalling and reactive oxygen species drive fibrosis and matrix remodelling in myxomatous mitral valves.Cardiovasc Res. 2013 Jul 1;99(1):175-84. doi: 10.1093/cvr/cvt083. Epub 2013 Apr 3. Cardiovasc Res. 2013. PMID: 23554457 Free PMC article.
-
TGFβ2 and TGFβ3 mediate appropriate context-dependent phenotype of rat valvular interstitial cells.iScience. 2021 Feb 3;24(3):102133. doi: 10.1016/j.isci.2021.102133. eCollection 2021 Mar 19. iScience. 2021. PMID: 33665554 Free PMC article.
-
The effect of physiological stretch and the valvular endothelium on mitral valve proteomes.Exp Biol Med (Maywood). 2019 Mar;244(3):241-251. doi: 10.1177/1535370219829006. Epub 2019 Feb 5. Exp Biol Med (Maywood). 2019. PMID: 30722697 Free PMC article.
-
Sex-related differences in gene expression by porcine aortic valvular interstitial cells.PLoS One. 2012;7(7):e39980. doi: 10.1371/journal.pone.0039980. Epub 2012 Jul 10. PLoS One. 2012. PMID: 22808080 Free PMC article.
References
-
- Lester W., Rosenthal A., Granton B., Gotlieb A.I. Porcine mitral valve interstitial cells in culture. Lab Invest. 1988;59:710–719. - PubMed
-
- Mulholland D.L., Gotlieb A.I. Cell biology of valvular interstitial cells. Can J Cardiol. 1996;12:231–236. - PubMed
-
- Filip D.A., Radu A., Simionescu M. Interstitial cells of the heart valves possess characteristics similar to smooth muscle cells. Circ Res. 1986;59:310–320. - PubMed
-
- Taylor P.M., Batten P., Brand N.J., Thomas P.S., Yacoub M.H. The cardiac valve interstitial cell. Int J Biochem Cell Biol. 2003;35:113–118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
