

Exploration of the copper-related compensatory response in the Belgrade rat model of genetic iron deficiency
- PMID: 21852364
- PMCID: PMC3220320
- DOI: 10.1152/ajpgi.00261.2011
Exploration of the copper-related compensatory response in the Belgrade rat model of genetic iron deficiency
Abstract
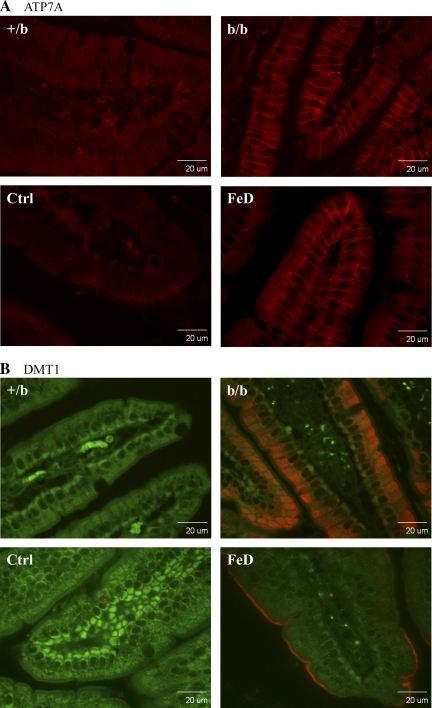
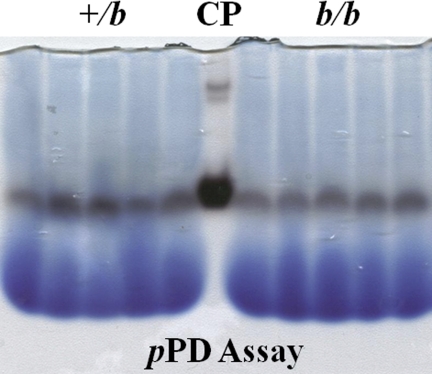
The Menkes copper ATPase (Atp7a) and metallothionein (Mt1a) are induced in the duodenum of iron-deficient rats, and serum and hepatic copper levels increase. Induction of a multi-copper ferroxidase (ceruloplasmin; Cp) has also been documented. These findings hint at an important role for Cu during iron deficiency. The intestinal divalent metal transporter 1 (Dmt1) is also induced during iron deficiency. The hypothesis that Dmt1 is involved in the copper-related compensatory response during iron deficiency was tested, utilizing a mutant Dmt1 rat model, namely the Belgrade (b/b) rat. Data from b/b rats were compared with phenotypically normal, heterozygous +/b rats. Intestinal Atp7a and Dmt1 expression was increased in b/b rats, whereas Mt1a expression was unchanged. Serum and liver copper levels did not increase in the Belgrades nor did Cp protein or activity. The lack of fully functional Dmt1 may thus partially blunt the compensatory response to iron deficiency by 1) decreasing copper levels in enterocytes, as exemplified by a lack of Mt1a induction and a lesser induction of Atp7a, 2) abolishing the frequently described increase in liver and serum copper, and 3) attenuating the documented increase in Cp expression and activity.
Figures
References
-
- Anderson GJ, Frazer DM, McKie AT, Vulpe CD. The ceruloplasmin homolog hephaestin and the control of intestinal iron absorption. Blood Cells Mol Dis 29: 367–375, 2002 - PubMed
-
- Arredondo M, Munoz P, Mura CV, Nunez MT. DMT1, a physiologically relevant apical Cu1+ transporter of intestinal cells. Am J Physiol Cell Physiol 284: C1525–C1530, 2003 - PubMed
-
- Bremner I. Involvement of metallothionein in the hepatic metabolism of copper. J Nutr 117: 19–29, 1987 - PubMed
-
- Canonne-Hergaux F, Fleming MD, Levy JE, Gauthier S, Ralph T, Picard V, Andrews NC, Gros P. The Nramp2/DMT1 iron transporter is induced in the duodenum of microcytic anemia mk mice but is not properly targeted to the intestinal brush border. Blood 96: 3964–3970, 2000 - PubMed
-
- Cartwright GE, Huguley CM, Jr, Aschenbrucker H, Fay J, Wintrobe MM. Studies on free erythrocyte protoporphyrin, plasma iron and plasma copper in normal and anemic subjects. Blood 3: 501–525, 1948. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous