

Mutational inactivation of STAG2 causes aneuploidy in human cancer
- PMID: 21852505
- PMCID: PMC3374335
- DOI: 10.1126/science.1203619
Mutational inactivation of STAG2 causes aneuploidy in human cancer
Abstract
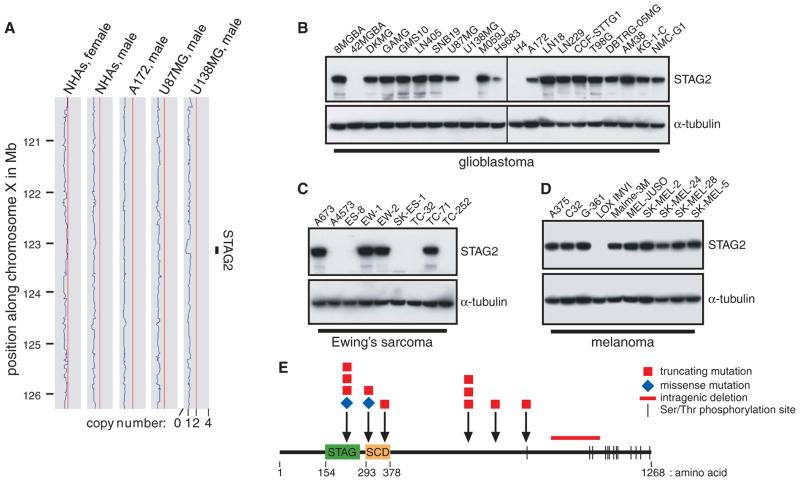
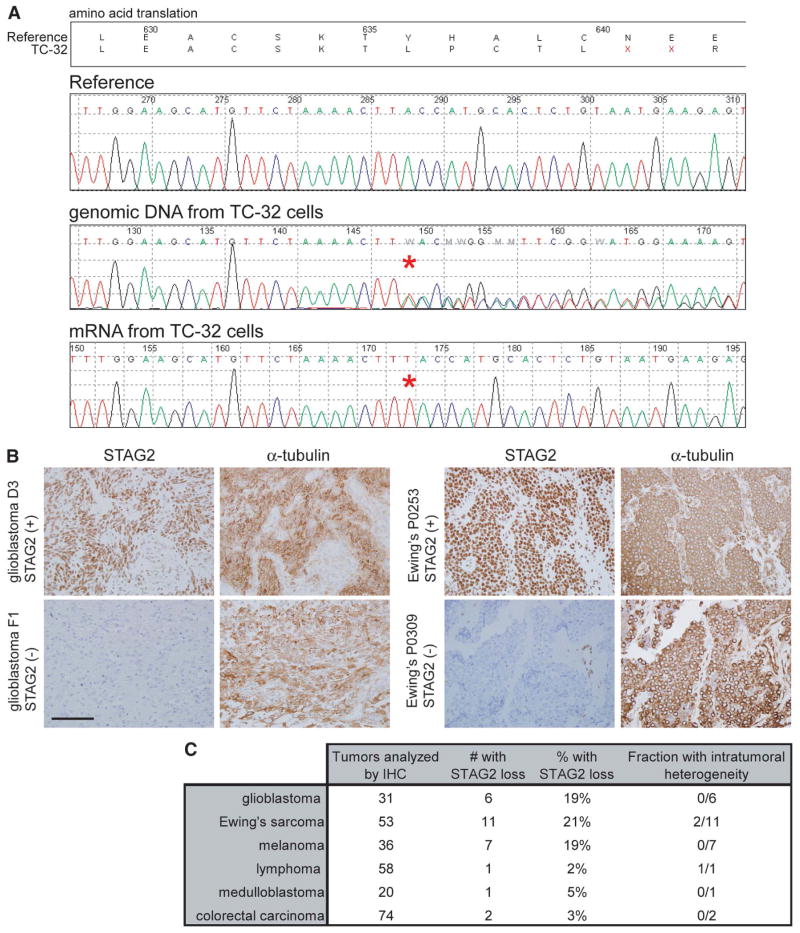
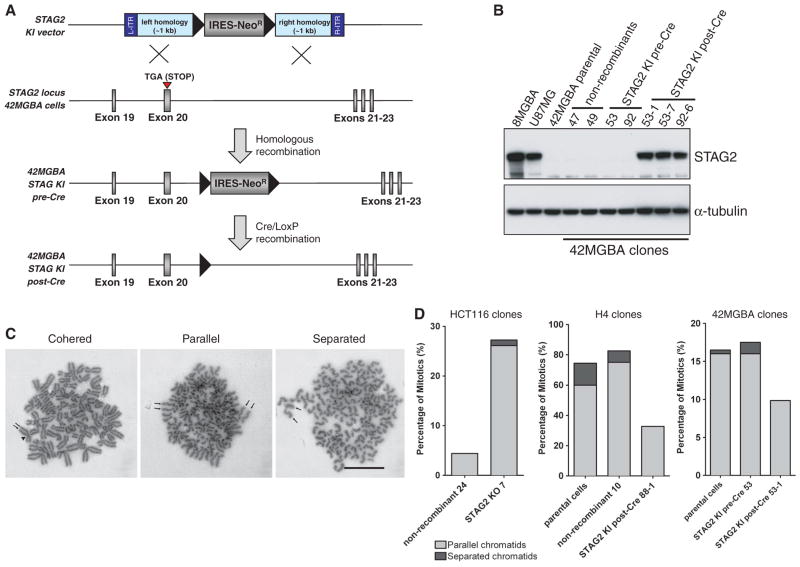
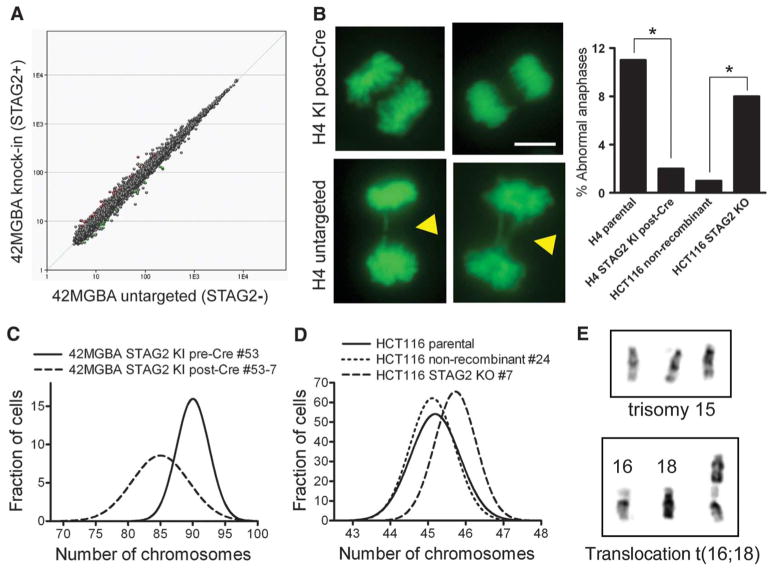
Most cancer cells are characterized by aneuploidy, an abnormal number of chromosomes. We have identified a clue to the mechanistic origins of aneuploidy through integrative genomic analyses of human tumors. A diverse range of tumor types were found to harbor deletions or inactivating mutations of STAG2, a gene encoding a subunit of the cohesin complex, which regulates the separation of sister chromatids during cell division. Because STAG2 is on the X chromosome, its inactivation requires only a single mutational event. Studying a near-diploid human cell line with a stable karyotype, we found that targeted inactivation of STAG2 led to chromatid cohesion defects and aneuploidy, whereas in two aneuploid human glioblastoma cell lines, targeted correction of the endogenous mutant alleles of STAG2 led to enhanced chromosomal stability. Thus, genetic disruption of cohesin is a cause of aneuploidy in human cancer.
Figures
Comment in
-
Cancer. Aneuploidy drives a mutator phenotype in cancer.Science. 2011 Aug 19;333(6045):942-3. doi: 10.1126/science.1211154. Science. 2011. PMID: 21852477 Free PMC article. No abstract available.
-
Aneuploidy stokes the fire.Nat Rev Genet. 2011 Aug 31;12(10):666. doi: 10.1038/nrg3070. Nat Rev Genet. 2011. PMID: 21878964 No abstract available.
-
A new mutation with staggering effects.Pigment Cell Melanoma Res. 2011 Dec;24(6):1081-2. doi: 10.1111/j.1755-148X.2011.00928.x. Pigment Cell Melanoma Res. 2011. PMID: 22216441 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
