

Nedd4-2 modulates renal Na+-Cl- cotransporter via the aldosterone-SGK1-Nedd4-2 pathway
- PMID: 21852580
- PMCID: PMC3171941
- DOI: 10.1681/ASN.2011020132
Nedd4-2 modulates renal Na+-Cl- cotransporter via the aldosterone-SGK1-Nedd4-2 pathway
Abstract
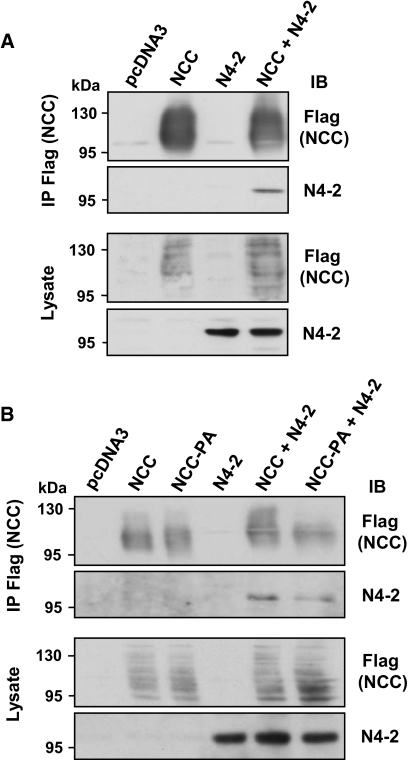
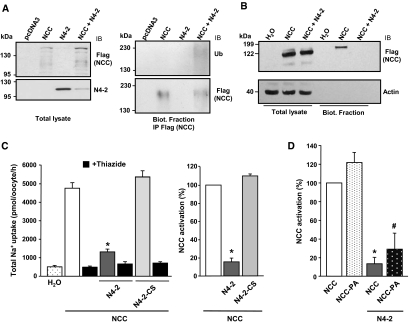
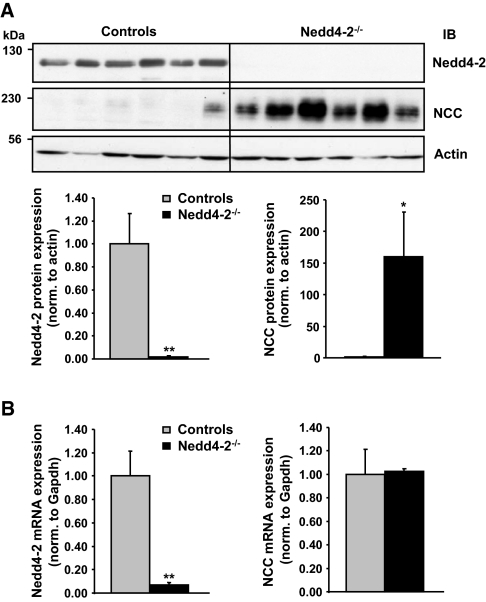
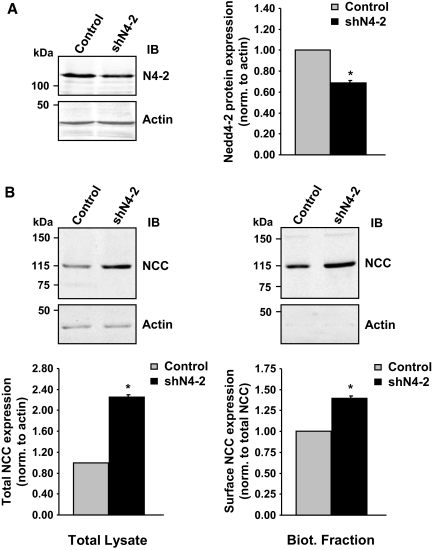
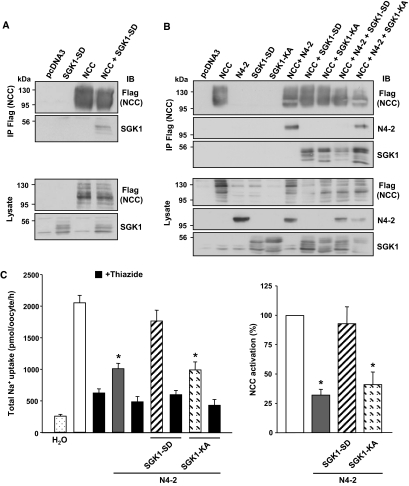
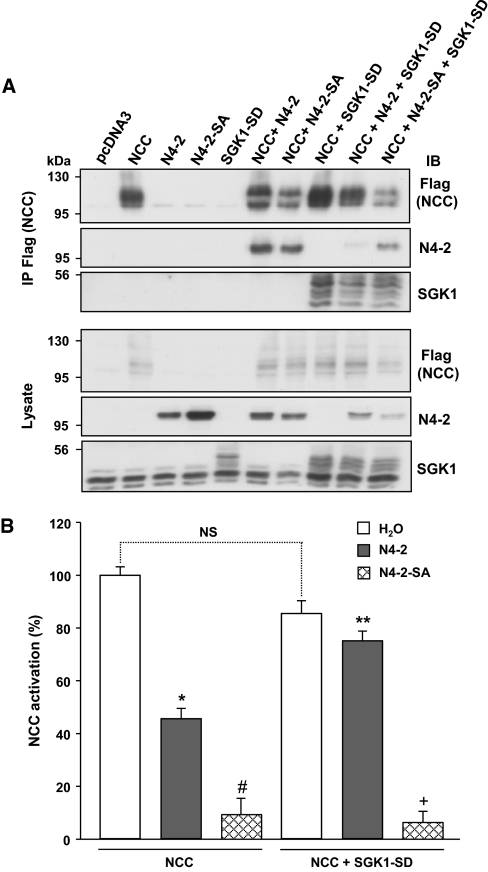
Regulation of renal Na(+) transport is essential for controlling blood pressure, as well as Na(+) and K(+) homeostasis. Aldosterone stimulates Na(+) reabsorption by the Na(+)-Cl(-) cotransporter (NCC) in the distal convoluted tubule (DCT) and by the epithelial Na(+) channel (ENaC) in the late DCT, connecting tubule, and collecting duct. Aldosterone increases ENaC expression by inhibiting the channel's ubiquitylation and degradation; aldosterone promotes serum-glucocorticoid-regulated kinase SGK1-mediated phosphorylation of the ubiquitin-protein ligase Nedd4-2 on serine 328, which prevents the Nedd4-2/ENaC interaction. It is important to note that aldosterone increases NCC protein expression by an unknown post-translational mechanism. Here, we present evidence that Nedd4-2 coimmunoprecipitated with NCC and stimulated NCC ubiquitylation at the surface of transfected HEK293 cells. In Xenopus laevis oocytes, coexpression of NCC with wild-type Nedd4-2, but not its catalytically inactive mutant, strongly decreased NCC activity and surface expression. SGK1 prevented this inhibition in a kinase-dependent manner. Furthermore, deficiency of Nedd4-2 in the renal tubules of mice and in cultured mDCT(15) cells upregulated NCC. In contrast to ENaC, Nedd4-2-mediated inhibition of NCC did not require the PY-like motif of NCC. Moreover, the mutation of Nedd4-2 at either serine 328 or 222 did not affect SGK1 action, and mutation at both sites enhanced Nedd4-2 activity and abolished SGK1-dependent inhibition. Taken together, these results suggest that aldosterone modulates NCC protein expression via a pathway involving SGK1 and Nedd4-2 and provides an explanation for the well-known aldosterone-induced increase in NCC protein expression.
Figures
References
-
- Loffing J, Loffing-Cueni D, Valderrabano V, Klausli L, Hebert SC, Rossier BC, Hoenderop JG, Bindels RJ, Kaissling B: Distribution of transcellular calcium and sodium transport pathways along mouse distal nephron. Am J Physiol Renal Physiol 281: F1021–F1027, 2001 - PubMed
-
- Bostanjoglo M, Reeves WB, Reilly RF, Velazquez H, Robertson N, Litwack G, Morsing P, Dorup J, Bachmann S, Ellison DH, Bostonjoglo M: 11Beta-hydroxysteroid dehydrogenase, mineralocorticoid receptor, and thiazide-sensitive Na-Cl cotransporter expression by distal tubules. J Am Soc Nephrol 9: 1347–1358, 1998 - PubMed
-
- Reilly RF, Ellison DH: Mammalian distal tubule: Physiology, pathophysiology, and molecular anatomy. Physiol Rev 80: 277–313, 2000 - PubMed
-
- Rusvai E, Naray-Fejes-Toth A: A new isoform of 11 beta-hydroxysteroid dehydrogenase in aldosterone target cells. J Biol Chem 268: 10717–10720, 1993 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
