

Structure and dynamics of the membrane-bound cytochrome P450 2C9
- PMID: 21852944
- PMCID: PMC3154944
- DOI: 10.1371/journal.pcbi.1002152
Structure and dynamics of the membrane-bound cytochrome P450 2C9
Abstract
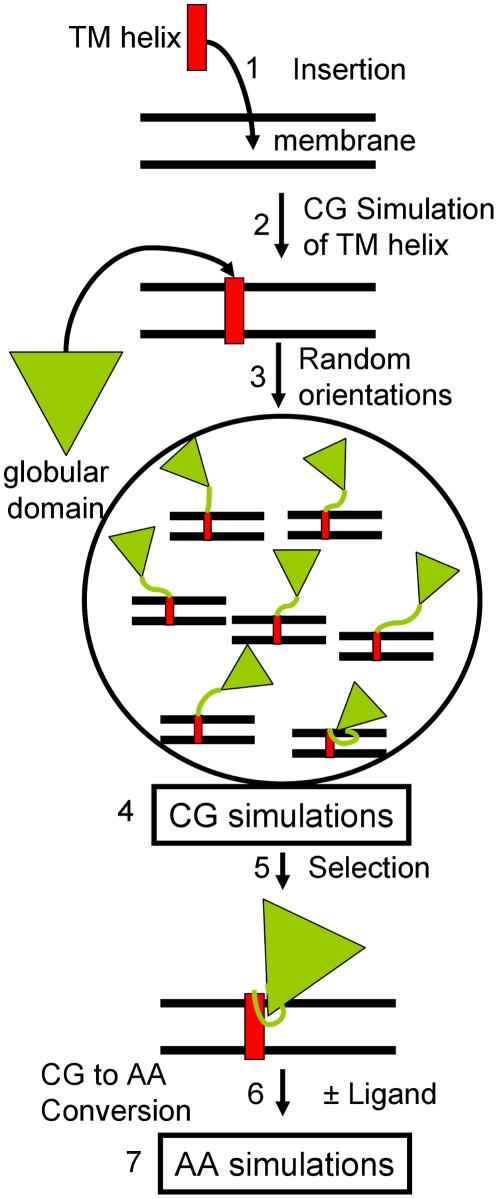
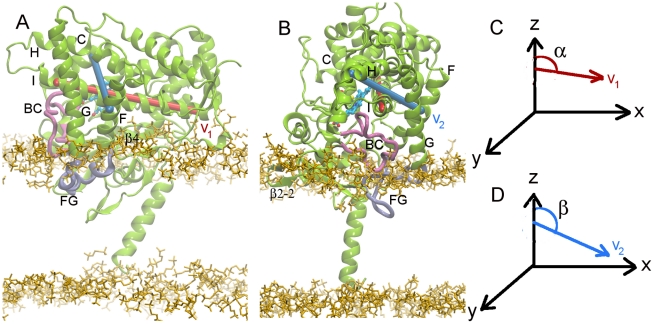
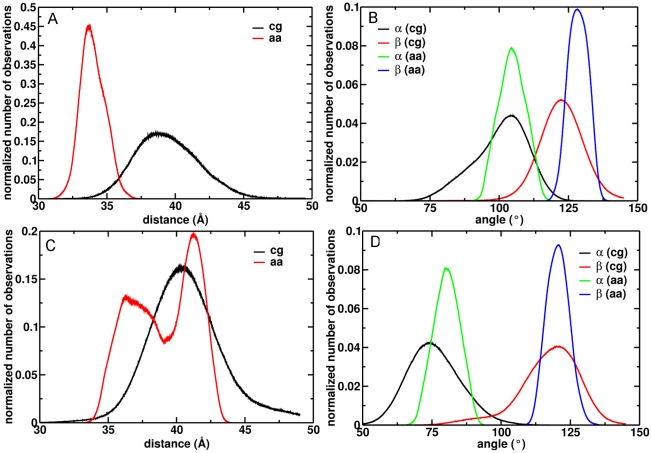
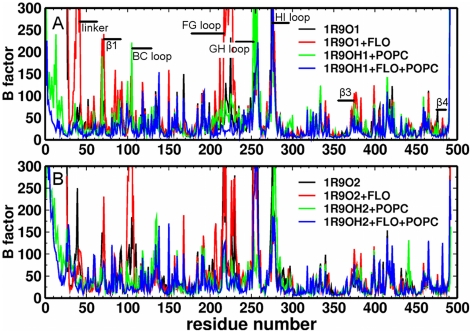
The microsomal, membrane-bound, human cytochrome P450 (CYP) 2C9 is a liver-specific monooxygenase essential for drug metabolism. CYPs require electron transfer from the membrane-bound CYP reductase (CPR) for catalysis. The structural details and functional relevance of the CYP-membrane interaction are not understood. From multiple coarse grained molecular simulations started with arbitrary configurations of protein-membrane complexes, we found two predominant orientations of CYP2C9 in the membrane, both consistent with experiments and conserved in atomic-resolution simulations. The dynamics of membrane-bound and soluble CYP2C9 revealed correlations between opening and closing of different tunnels from the enzyme's buried active site. The membrane facilitated the opening of a tunnel leading into it by stabilizing the open state of an internal aromatic gate. Other tunnels opened selectively in the simulations of product-bound CYP2C9. We propose that the membrane promotes binding of liposoluble substrates by stabilizing protein conformations with an open access tunnel and provide evidence for selective substrate access and product release routes in mammalian CYPs. The models derived here are suitable for extension to incorporate other CYPs for oligomerization studies or the CYP reductase for studies of the electron transfer mechanism, whereas the modeling procedure is generally applicable to study proteins anchored in the bilayer by a single transmembrane helix.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
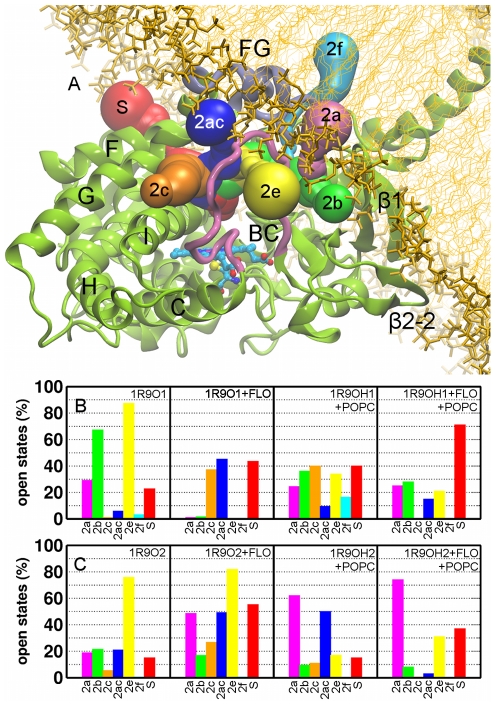
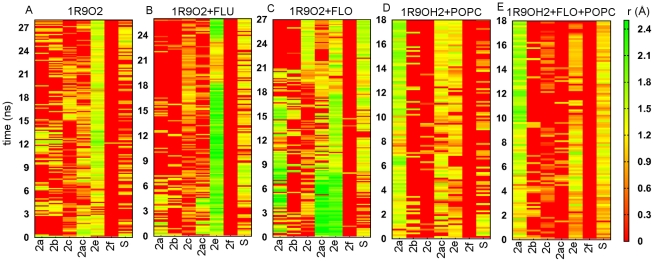
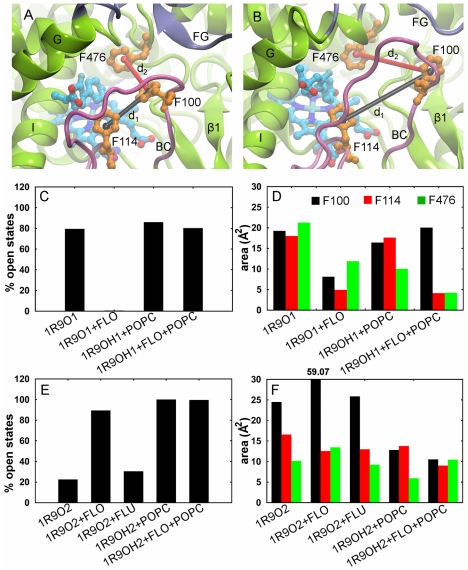
References
-
- Guengerich FP. Cytochrome P450 and chemical toxicology. Chem Res Toxicol. 2008;21:70–83. - PubMed
-
- Black SD, Martin ST, Smith CA. Membrane topology of liver microsomal cytochrome P450 2B4 determined via monoclonal antibodies directed to the halt-transfer signal. Biochemistry. 1994;33:6945–6951. - PubMed
-
- Strobel HW, Lu AY, Heidema J, Coon MJ. Phosphatidylcholine requirement in the enzymatic reduction of hemoprotein P-450 and in fatty acid, hydrocarbon, and drug hydroxylation. J Biol Chem. 1970;245:4851–4854. - PubMed
-
- Nath A, Grinkova YV, Sligar SG, Atkins WM. Ligand binding to cytochrome P450 3A4 in phospholipid bilayer nanodiscs: the effect of model membranes. J Biol Chem. 2007;282:28309–28320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BBS/B/16011/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BEP17032/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- B19456/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT083547MA/WT_/Wellcome Trust/United Kingdom
- BB/H000267/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
