

Multiple nuclear gene phylogenetic analysis of the evolution of dioecy and sex chromosomes in the genus Silene
- PMID: 21853022
- PMCID: PMC3154253
- DOI: 10.1371/journal.pone.0021915
Multiple nuclear gene phylogenetic analysis of the evolution of dioecy and sex chromosomes in the genus Silene
Abstract
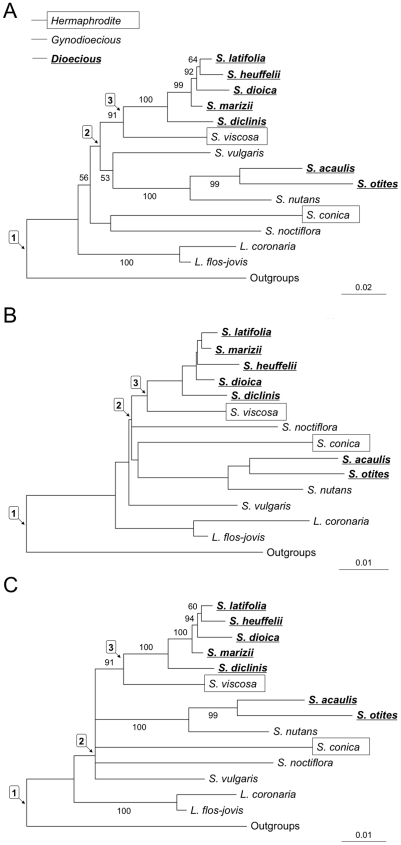
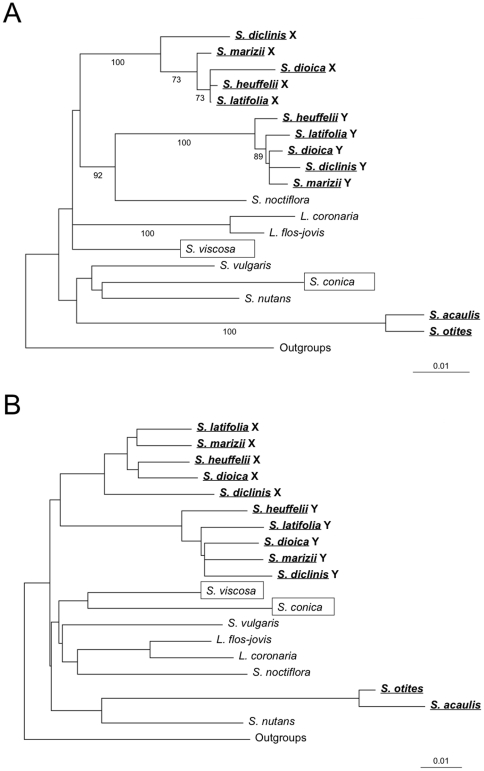
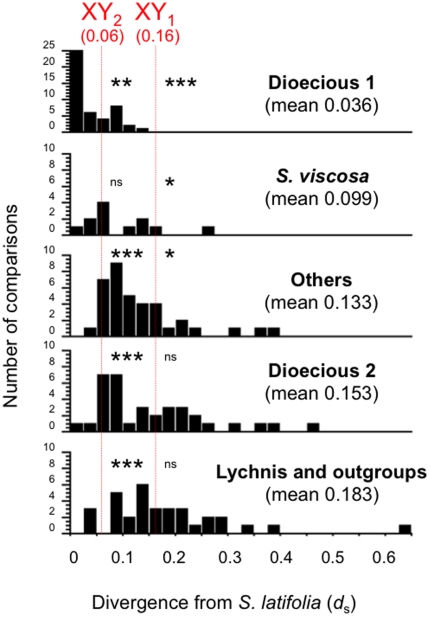
In the plant genus Silene, separate sexes and sex chromosomes are believed to have evolved twice. Silene species that are wholly or largely hermaphroditic are assumed to represent the ancestral state from which dioecy evolved. This assumption is important for choice of outgroup species for inferring the genetic and chromosomal changes involved in the evolution of dioecy, but is mainly based on data from a single locus (ITS). To establish the order of events more clearly, and inform outgroup choice, we therefore carried out (i) multi-nuclear-gene phylogenetic analyses of 14 Silene species (including 7 hermaphrodite or gynodioecious species), representing species from both Silene clades with dioecious members, plus a more distantly related outgroup, and (ii) a BayesTraits character analysis of the evolution of dioecy. We confirm two origins of dioecy within this genus in agreement with recent work on comparing sex chromosomes from both clades with dioecious species. We conclude that sex chromosomes evolved after the origin of Silene and within a clade that includes only S. latifolia and its closest relatives. We estimate that sex chromosomes emerged soon after the split with the ancestor of S. viscosa, the probable closest non-dioecious S. latifolia relative among the species included in our study.
Conflict of interest statement
Figures
Similar articles
-
Evolution of sex determination systems with heterogametic males and females in silene.Evolution. 2013 Dec;67(12):3669-77. doi: 10.1111/evo.12223. Epub 2013 Sep 2. Evolution. 2013. PMID: 24299418
-
Independent origin of sex chromosomes in two species of the genus Silene.Genetics. 2008 Jun;179(2):1129-33. doi: 10.1534/genetics.107.085670. Genetics. 2008. PMID: 18558658 Free PMC article.
-
The genomics of plant sex chromosomes.Plant Sci. 2015 Jul;236:126-35. doi: 10.1016/j.plantsci.2015.03.019. Epub 2015 Apr 2. Plant Sci. 2015. PMID: 26025526 Review.
-
Evolution of reproductive systems in the genus Silene.Proc Biol Sci. 1996 Apr 22;263(1369):409-14. doi: 10.1098/rspb.1996.0062. Proc Biol Sci. 1996. PMID: 18386410
-
Silene latifolia: the classical model to study heteromorphic sex chromosomes.Cytogenet Genome Res. 2010 Jul;129(1-3):250-62. doi: 10.1159/000314285. Epub 2010 Jun 10. Cytogenet Genome Res. 2010. PMID: 20551610 Review.
Cited by
-
SEX-DETector: A Probabilistic Approach to Study Sex Chromosomes in Non-Model Organisms.Genome Biol Evol. 2016 Aug 29;8(8):2530-43. doi: 10.1093/gbe/evw172. Genome Biol Evol. 2016. PMID: 27492231 Free PMC article.
-
Has adaptation occurred in males and females since separate sexes evolved in the plant Silene latifolia?Proc Biol Sci. 2018 Jul 25;285(1883):20172824. doi: 10.1098/rspb.2017.2824. Proc Biol Sci. 2018. PMID: 30051860 Free PMC article.
-
Identification of white campion (Silene latifolia) guaiacol O-methyltransferase involved in the biosynthesis of veratrole, a key volatile for pollinator attraction.BMC Plant Biol. 2012 Aug 31;12:158. doi: 10.1186/1471-2229-12-158. BMC Plant Biol. 2012. PMID: 22937972 Free PMC article.
-
Identification of internal reference genes for gene expression normalization between the two sexes in dioecious white Campion.PLoS One. 2014 Mar 27;9(3):e92893. doi: 10.1371/journal.pone.0092893. eCollection 2014. PLoS One. 2014. PMID: 24675788 Free PMC article.
-
An angiosperm-wide analysis of the gynodioecy-dioecy pathway.Ann Bot. 2014 Sep;114(3):539-48. doi: 10.1093/aob/mcu134. Epub 2014 Aug 4. Ann Bot. 2014. PMID: 25091207 Free PMC article.
References
-
- Charlesworth D, Guttman DS. The evolution of dioecy and plant sex chromosome systems. In: Ainsworth CCe., editor. Sex Determination in Plants. Oxford: 1999. pp. 25–49.
-
- Liu Z, Moore PH, Ma H, Ackerman CM, Ragiba M, et al. A primitive Y chromosome in Papaya marks the beginning of sex chromosome evolution. Nature. 2004;427:348–352. - PubMed
-
- Spigler R, Lewers K, Main D, Ashman T-L. Genetic mapping of sex determination in a wild strawberry, Fragaria virginiana, reveals earliest form of sex chromosome. Heredity. 2008;101:507–517. - PubMed
-
- Desfeux C, Maurice S, Henry JP, Lejeune B, Gouyon PH. Evolution of reproductive systems in the genus Silene. Proc Biol Sci. 1996;263:409–414. - PubMed
-
- Eggens F, Popp M, Nepokroeff M, Wagner W, Oxelman B. The origin and number of introductions of the Hawaiian endemic Silene species (Caryophyllaceae). American Journal of Botany. 2007;94:210–218. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
