

Single molecule analysis of c-myb alternative splicing reveals novel classifiers for precursor B-ALL
- PMID: 21853052
- PMCID: PMC3154906
- DOI: 10.1371/journal.pone.0022880
Single molecule analysis of c-myb alternative splicing reveals novel classifiers for precursor B-ALL
Abstract
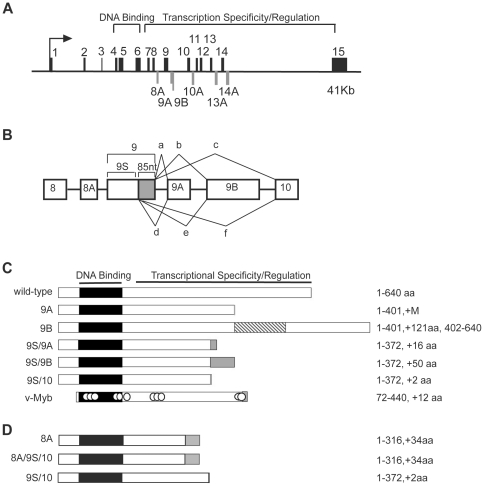
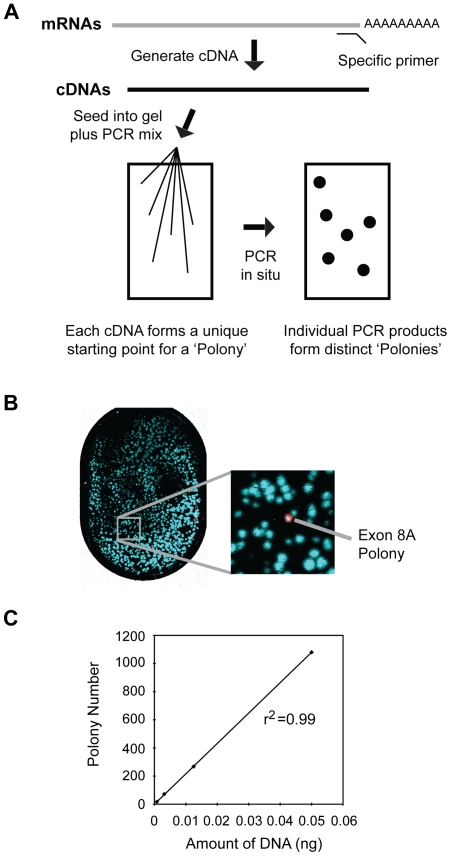
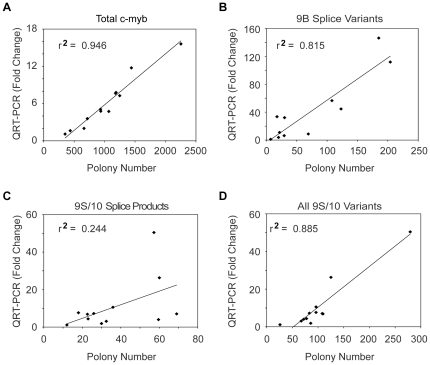
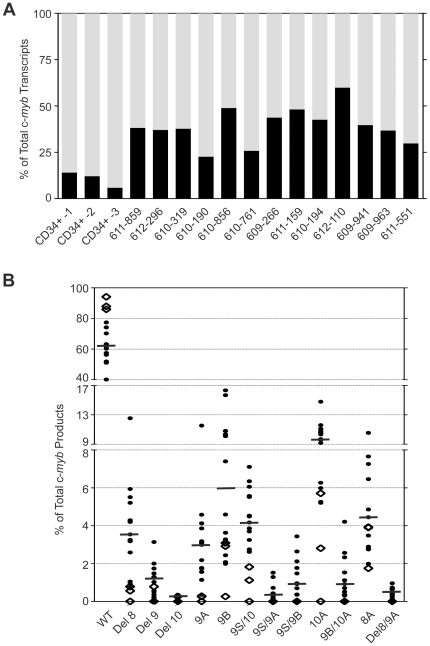
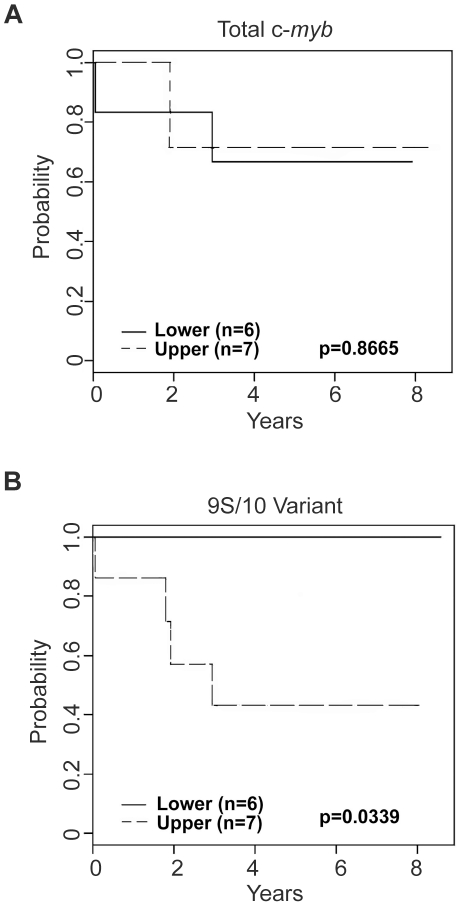
The c-Myb transcription factor, a key regulator of proliferation and differentiation in hematopoietic and other cell types, has an N-terminal DNA binding domain and a large C-terminal domain responsible for transcriptional activation, negative regulation and determining target gene specificity. Overexpression and rearrangement of the c-myb gene (MYB) has been reported in some patients with leukemias and other types of cancers, implicating activated alleles of c-myb in the development of human tumors. Alternative RNA splicing can produce variants of c-myb with qualitatively distinct transcriptional activities that may be involved in transformation and leukemogenesis. Here, by performing a detailed, single molecule assay we found that c-myb alternative RNA splicing was elevated and much more complex in leukemia samples than in cell lines or CD34+ hematopoietic progenitor cells from normal donors. The results revealed that leukemia samples express more than 60 different c-myb splice variants, most of which have multiple alternative splicing events and were not detectable by conventional microarray or PCR approaches. For example, the single molecule assay detected 21 and 22 splice variants containing the 9B and 9S exons, respectively, most of which encoded unexpected variant forms of c-Myb protein. Furthermore, the detailed analysis identified some splice variants whose expression correlated with poor survival in a small cohort of precursor B-ALL samples. Our findings indicate that single molecule assays can reveal complexities in c-myb alternative splicing that have potential as novel biomarkers and could help explain the role of c-Myb variants in the development of human leukemia.
Conflict of interest statement
Figures
References
-
- Mucenski ML, McLain K, Kier AB, Swerdlow SH, Schreiner CM, et al. A functional c-myb gene is required for normal murine fetal hepatic hematopoiesis. Cell. 1991;65:677–689. - PubMed
-
- Sandberg ML, Sutton SE, Pletcher MT, Wiltshire T, Tarantino LM, et al. c-Myb and p300 regulate hematopoietic stem cell proliferation and differentiation. Dev Cell. 2005;8:153–166. - PubMed
-
- Sakamoto H, Dai G, Tsujino K, Hashimoto K, Huang X, et al. Proper levels of c-Myb are discretely defined at distinct steps of hematopoietic cell development. Blood. 2006;108:896–903. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
