

An atypical unfolded protein response in heat shocked cells
- PMID: 21853144
- PMCID: PMC3154502
- DOI: 10.1371/journal.pone.0023512
An atypical unfolded protein response in heat shocked cells
Abstract
Background: The heat shock response (HSR) and the unfolded protein response (UPR) are both activated by proteotoxic stress, although in different compartments, and share cellular resources. How these resources are allocated when both responses are active is not known. Insight in possible crosstalk will help understanding the consequences of failure of these systems in (age-related) disease.
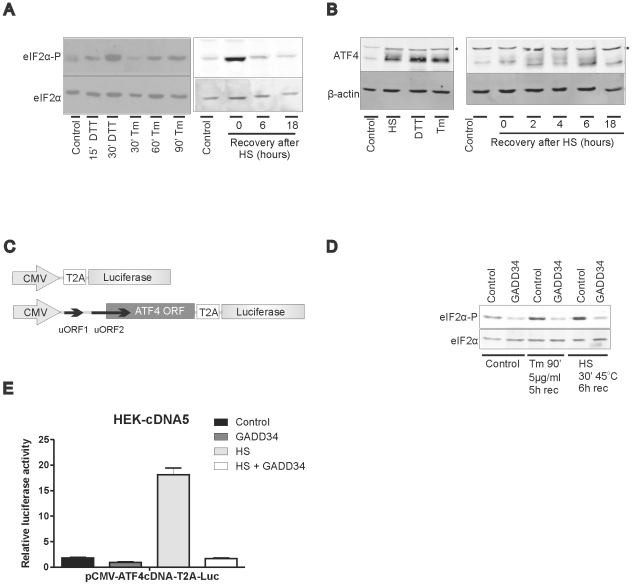
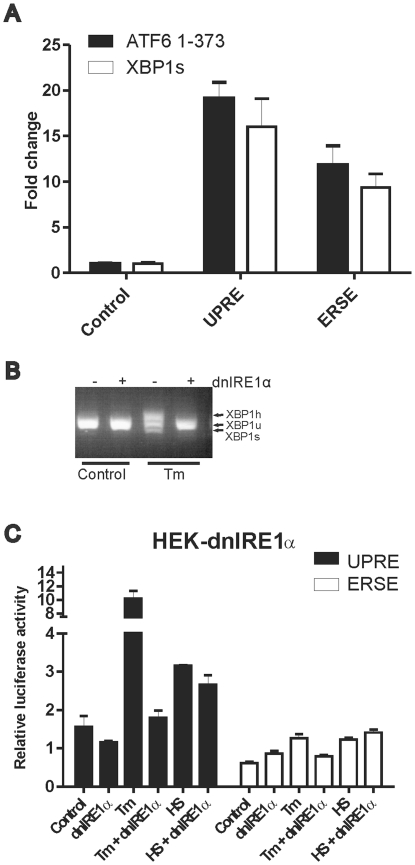
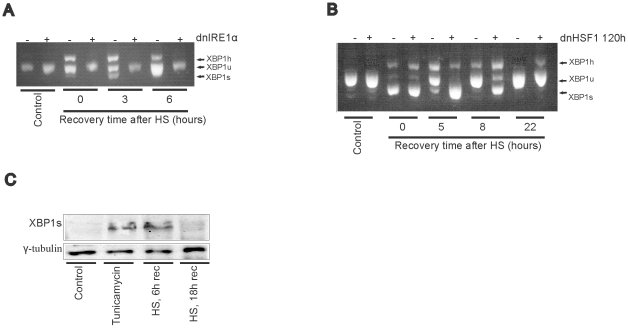
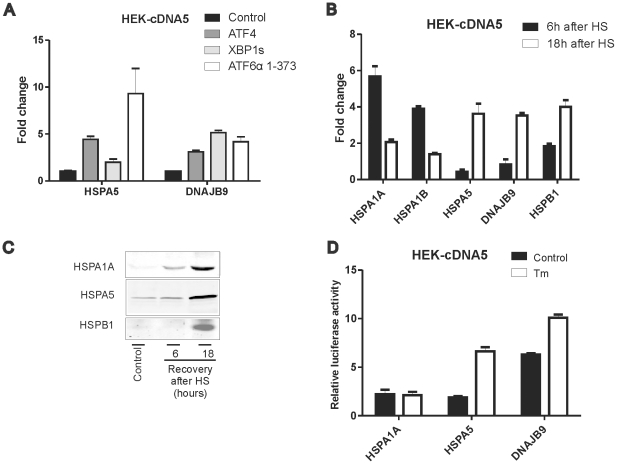
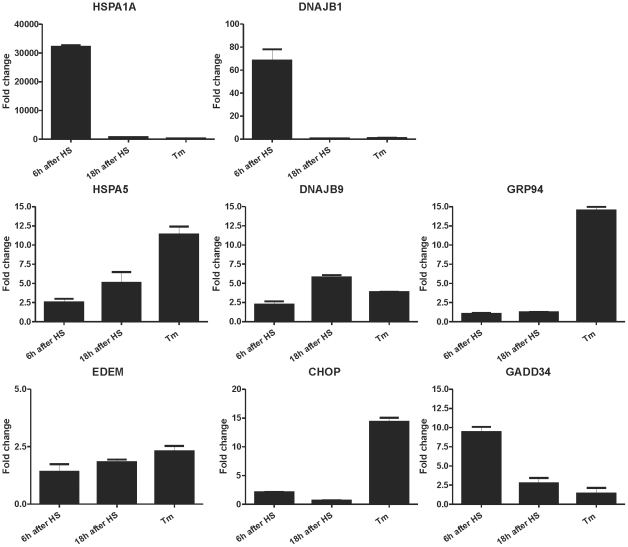
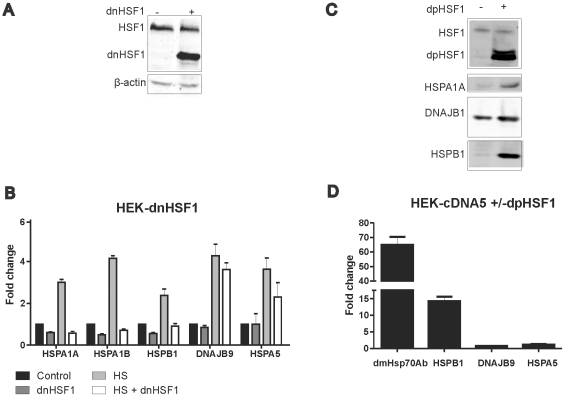
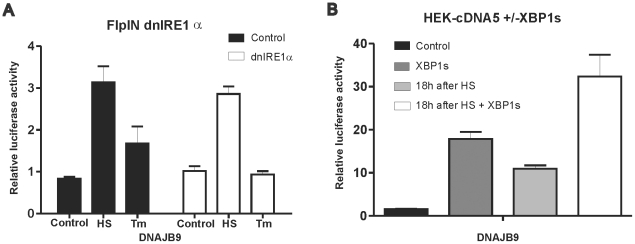
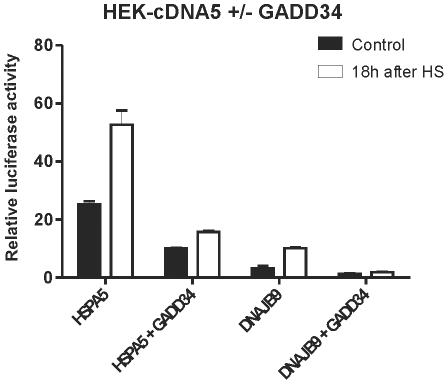
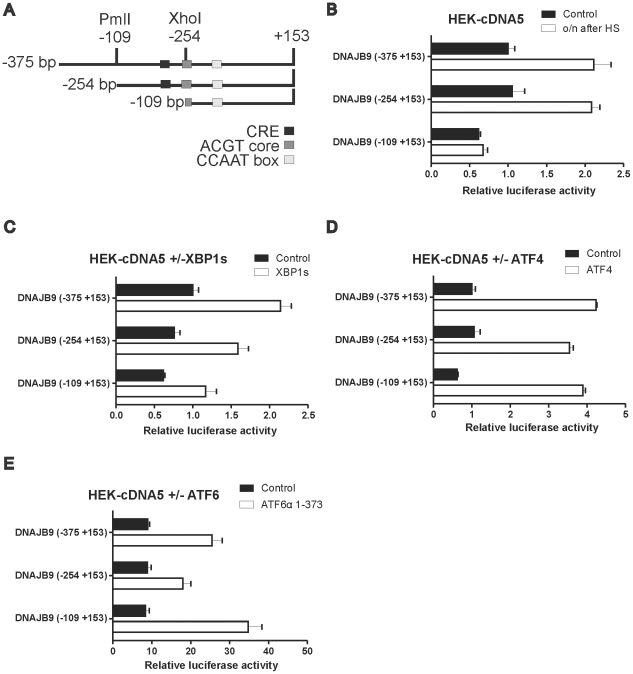
Results: In heat stressed HEK293 cells synthesis of the canonical UPR transcription factors XBP1s and ATF4 was detected as well as HSF1 independent activation of the promoters of the ER resident chaperones HSPA5 (BiP) and DNAJB9 (ERdj4). However, the heat stress activation of the DNAJB9 promoter, a XBP1s target, was not blocked in cells expressing a dominant negative IRE1α mutant, and thus did not require XBP1s. Furthermore, the DNA element required for heat stress activation of the DNAJB9 promoter is distinct from the ATF4 and ATF6 target elements; even though inhibition of eIF2α phosphorylation resulted in a decreased activation of the DNAJB9 promoter upon heat stress, suggesting a role for an eIF2α phosphorylation dependent product.
Conclusions: The initial step in the UPR, synthesis of transcription factors, is activated by heat stress but the second step, transcriptional transactivation by these factors, is blocked and these pathways of the UPR are thus not productive. Expression of canonical ER chaperones is part of the response of heat stressed cells but another set of transcription factors has been recruited to regulate expression of these ER chaperones.
Conflict of interest statement
Figures
Similar articles
-
Gene regulatory network of unfolded protein response genes in endoplasmic reticulum stress.Cell Stress Chaperones. 2013 Jan;18(1):11-23. doi: 10.1007/s12192-012-0351-5. Epub 2012 Jul 18. Cell Stress Chaperones. 2013. PMID: 22802018 Free PMC article.
-
A novel feedback loop regulates the response to endoplasmic reticulum stress via the cooperation of cytoplasmic splicing and mRNA translation.Mol Cell Biol. 2012 Mar;32(5):992-1003. doi: 10.1128/MCB.06665-11. Epub 2012 Jan 3. Mol Cell Biol. 2012. PMID: 22215619 Free PMC article.
-
XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response.Mol Cell Biol. 2003 Nov;23(21):7448-59. doi: 10.1128/MCB.23.21.7448-7459.2003. Mol Cell Biol. 2003. PMID: 14559994 Free PMC article.
-
Roles of heat shock factor 1 beyond the heat shock response.Cell Mol Life Sci. 2018 Aug;75(16):2897-2916. doi: 10.1007/s00018-018-2836-6. Epub 2018 May 17. Cell Mol Life Sci. 2018. PMID: 29774376 Free PMC article. Review.
-
Transcriptional reprogramming at the intersection of the heat shock response and proteostasis.Mol Cell. 2024 Jan 4;84(1):80-93. doi: 10.1016/j.molcel.2023.11.024. Epub 2023 Dec 15. Mol Cell. 2024. PMID: 38103561 Review.
Cited by
-
The aggregation-prone intracellular serpin SRP-2 fails to transit the ER in Caenorhabditis elegans.Genetics. 2015 May;200(1):207-19. doi: 10.1534/genetics.115.176180. Epub 2015 Mar 18. Genetics. 2015. PMID: 25786854 Free PMC article.
-
Heat shock protein 70 (Hsp70) mediates Zika virus entry, replication, and egress from host cells.Emerg Microbes Infect. 2019;8(1):8-16. doi: 10.1080/22221751.2018.1557988. Emerg Microbes Infect. 2019. PMID: 30866755 Free PMC article.
-
BAG3 induces the sequestration of proteasomal clients into cytoplasmic puncta: implications for a proteasome-to-autophagy switch.Autophagy. 2014 Sep;10(9):1603-21. doi: 10.4161/auto.29409. Epub 2014 Jul 10. Autophagy. 2014. PMID: 25046115 Free PMC article.
-
The heat-shock response co-inducer arimoclomol protects against retinal degeneration in rhodopsin retinitis pigmentosa.Cell Death Dis. 2014 May 22;5(5):e1236. doi: 10.1038/cddis.2014.214. Cell Death Dis. 2014. PMID: 24853414 Free PMC article.
-
The Arabidopsis J-Protein AtDjC5 Facilitates Thermotolerance Likely by Aiding in the ER Stress Response.Int J Mol Sci. 2022 Oct 28;23(21):13134. doi: 10.3390/ijms232113134. Int J Mol Sci. 2022. PMID: 36361922 Free PMC article.
References
-
- Zhang K, Kaufman RJ. The unfolded protein response: a stress signaling pathway critical for health and disease. Neurology. 2006;66:S102–109. - PubMed
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319:916–919. - PubMed
-
- Powers ET, Morimoto RI, Dillin A, Kelly JW, Balch WE. Biological and chemical approaches to diseases of proteostasis deficiency. Annu Rev Biochem. 2009;78:959–991. - PubMed
-
- Broadley SA, Hartl FU. The role of molecular chaperones in human misfolding diseases. FEBS Lett. 2009;583:2647–2653. - PubMed
-
- Morimoto RI. Regulation of the heat shock transcriptional response: cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Genes Dev. 1998;12:3788–3796. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
