

Commitment to a cellular transition precedes genome-wide transcriptional change
- PMID: 21855792
- PMCID: PMC3160620
- DOI: 10.1016/j.molcel.2011.06.024
Commitment to a cellular transition precedes genome-wide transcriptional change
Abstract
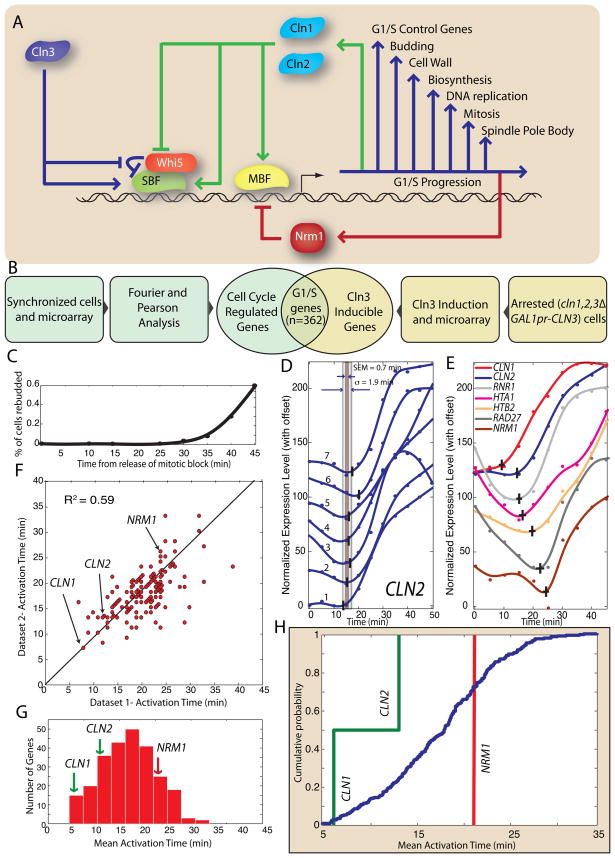
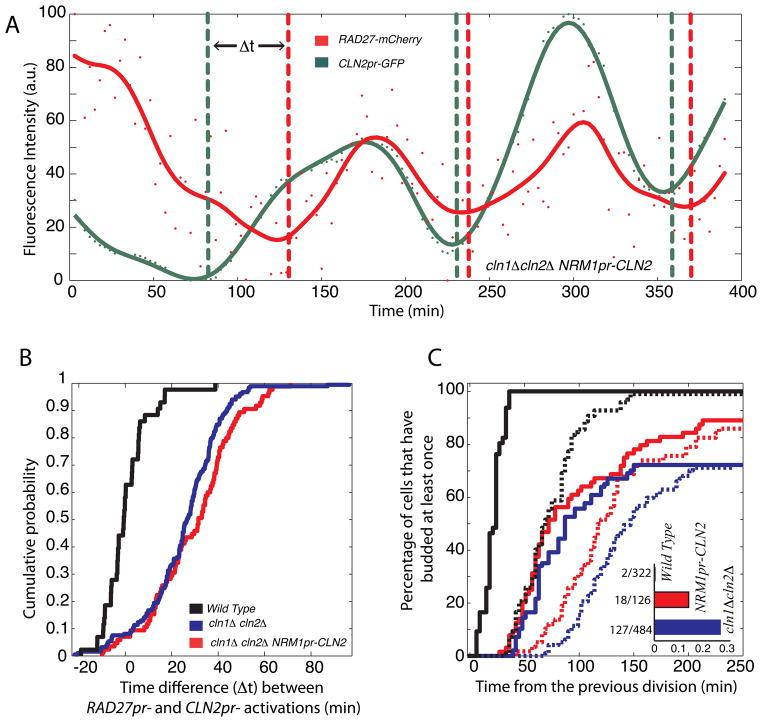
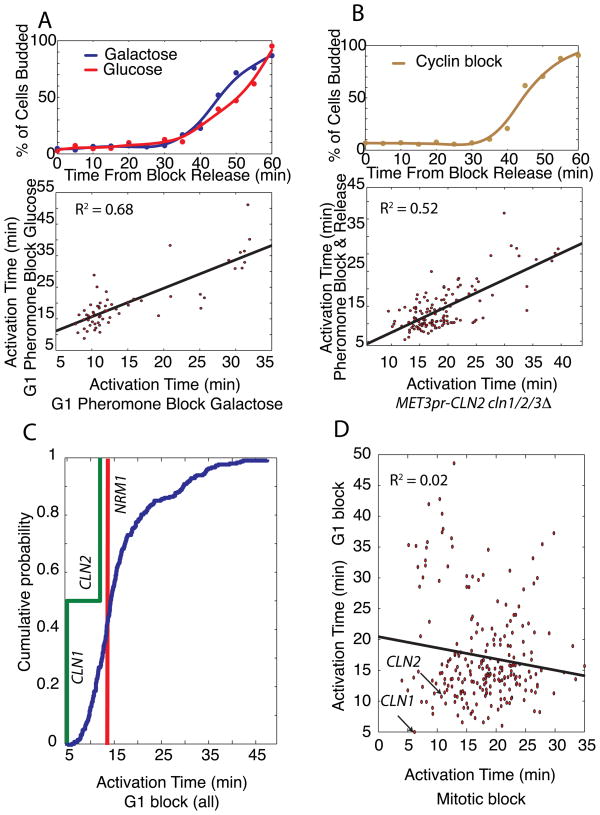
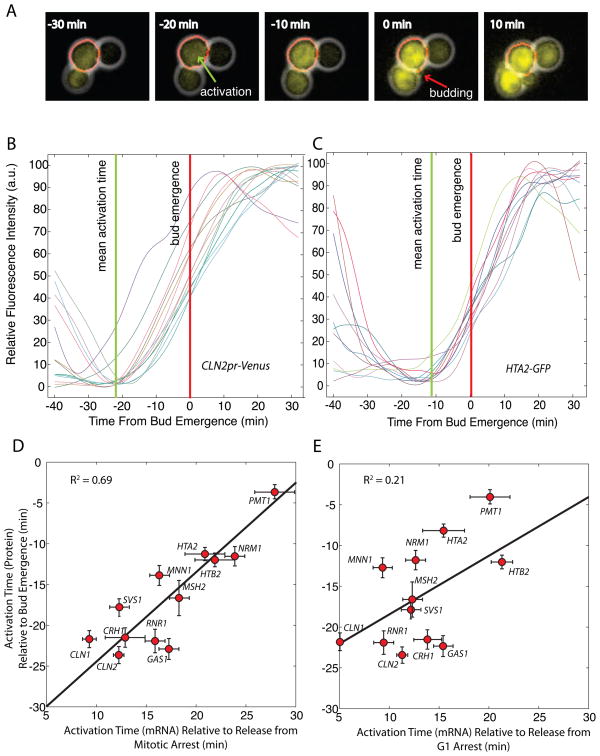
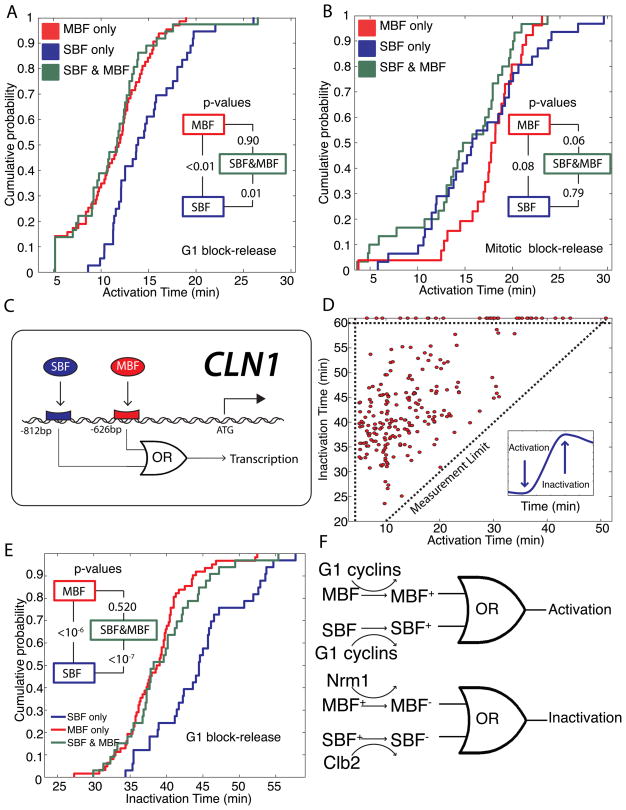
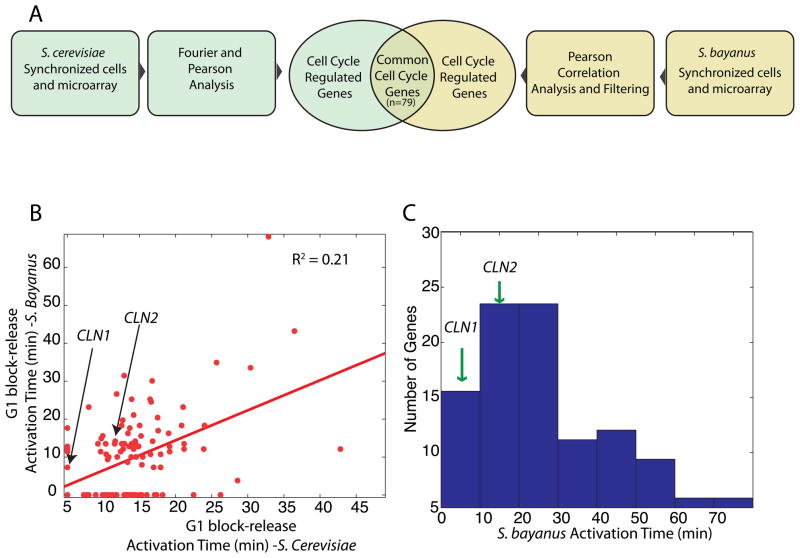
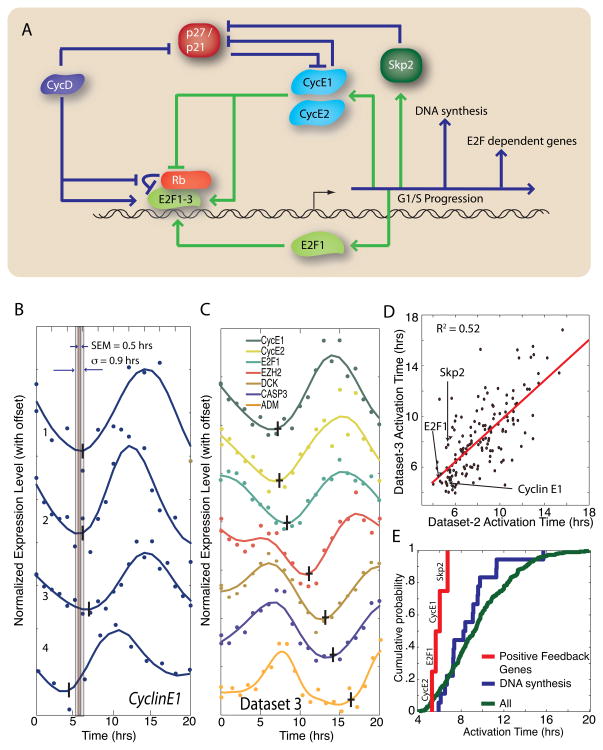
In budding yeast, commitment to cell division corresponds to activating the positive feedback loop of G1 cyclins controlled by the transcription factors SBF and MBF. This pair of transcription factors has over 200 targets, implying that cell-cycle commitment coincides with genome-wide changes in transcription. Here, we find that genes within this regulon have a well-defined distribution of transcriptional activation times. Combinatorial use of SBF and MBF results in a logical OR function for gene expression and partially explains activation timing. Activation of G1 cyclin expression precedes the activation of the bulk of the G1/S regulon, ensuring that commitment to cell division occurs before large-scale changes in transcription. Furthermore, we find similar positive feedback-first regulation in the yeasts S. bayanus and S. cerevisiae, as well as human cells. The widespread use of the feedback-first motif in eukaryotic cell-cycle control, implemented by nonorthologous proteins, suggests its frequent deployment at cellular transitions.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Simple rules for complex processes: new lessons from the budding yeast cell cycle.Mol Cell. 2011 Aug 19;43(4):497-500. doi: 10.1016/j.molcel.2011.08.002. Mol Cell. 2011. PMID: 21855788 Free PMC article.
References
-
- Amon A, Tyers M, et al. Mechanisms that help the yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins and repress G1 cyclins. Cell. 1993;74(6):993–1007. - PubMed
-
- Andrews BJ, Herskowitz I. Identification of a DNA binding factor involved in cell-cycle control of the yeast HO gene. Cell. 1989;57(1):21–9. - PubMed
-
- Bean JM, Siggia ED, et al. Coherence and timing of cell cycle Start examined at single-cell resolution. Mol Cell. 2006;21(1):3–14. - PubMed
-
- Blagosklonny MV, Pardee AB. The restriction point of the cell cycle. Cell Cycle. 2002;1(2):103–10. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
