

Ube2w and ataxin-3 coordinately regulate the ubiquitin ligase CHIP
- PMID: 21855799
- PMCID: PMC3166620
- DOI: 10.1016/j.molcel.2011.05.036
Ube2w and ataxin-3 coordinately regulate the ubiquitin ligase CHIP
Abstract
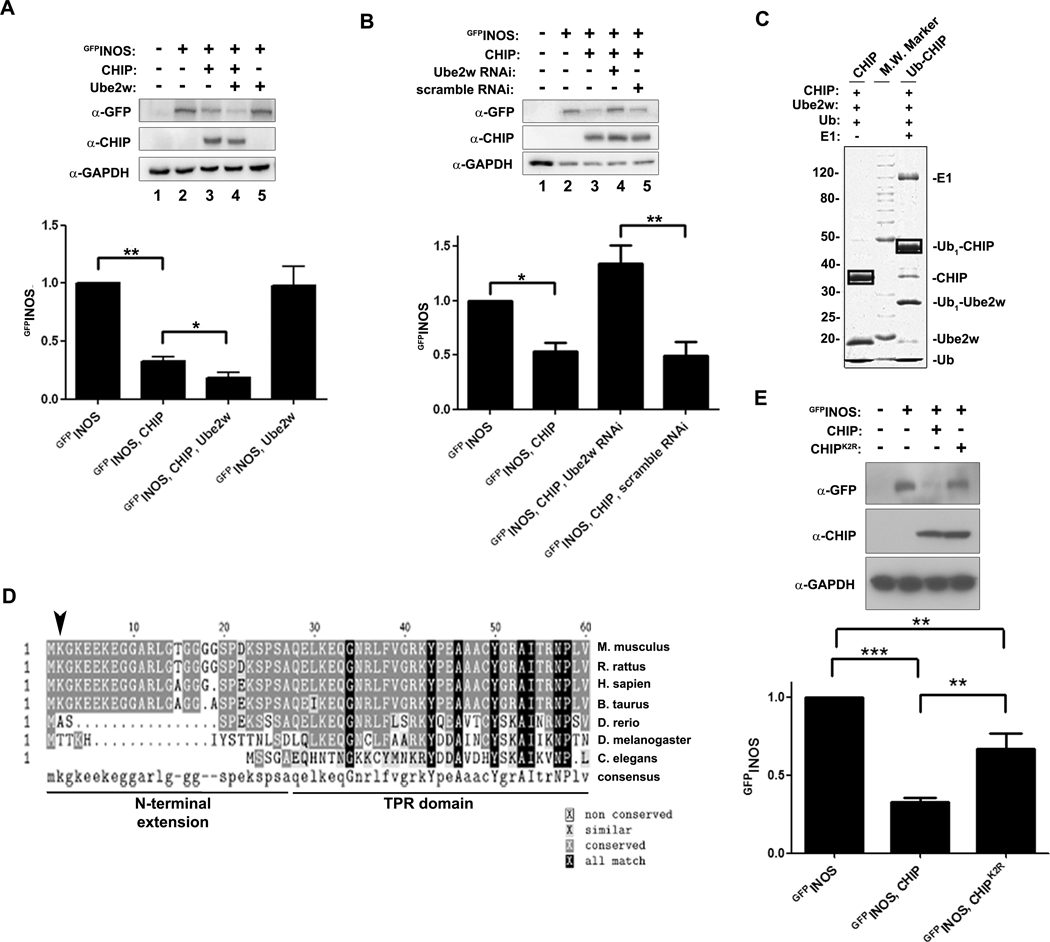
The mechanisms by which ubiquitin ligases are regulated remain poorly understood. Here we describe a series of molecular events that coordinately regulate CHIP, a neuroprotective E3 implicated in protein quality control. Through their opposing activities, the initiator E2, Ube2w, and the specialized deubiquitinating enzyme (DUB), ataxin-3, participate in initiating, regulating, and terminating the CHIP ubiquitination cycle. Monoubiquitination of CHIP by Ube2w stabilizes the interaction between CHIP and ataxin-3, which through its DUB activity limits the length of chains attached to CHIP substrates. Upon completion of substrate ubiquitination, ataxin-3 deubiquitinates CHIP, effectively terminating the reaction. Our results suggest that functional pairing of E3s with ataxin-3 or similar DUBs represents an important point of regulation in ubiquitin-dependent protein quality control. In addition, the results shed light on disease pathogenesis in SCA3, a neurodegenerative disorder caused by polyglutamine expansion in ataxin-3.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures






References
-
- Arndt V, Dick N, Tawo R, Dreiseidler M, Wenzel D, Hesse M, Furst DO, Saftig P, Saint R, Fleischmann BK, et al. Chaperone-assisted selective autophagy is essential for muscle maintenance. Curr Biol. 2010;20:143–148. - PubMed
-
- Burnett B, Li F, Pittman RN. The polyglutamine neurodegenerative protein ataxin-3 binds polyubiquitylated proteins and has ubiquitin protease activity. Hum Mol Genet. 2003;12:3195–3205. - PubMed
-
- Christensen DE, Brzovic PS, Klevit RE. E2-BRCA1 RING interactions dictate synthesis of mono- or specific polyubiquitin chain linkages. Nat Struct Mol Biol. 2007;14:941–948. - PubMed
-
- Connell P, Ballinger CA, Jiang J, Wu Y, Thompson LJ, Hohfeld J, Patterson C. The co-chaperone CHIP regulates protein triage decisions mediated by heat-shock proteins. Nat Cell Biol. 2001;3:93–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01AG034228/AG/NIA NIH HHS/United States
- K99 NS064097/NS/NINDS NIH HHS/United States
- R01NS038712/NS/NINDS NIH HHS/United States
- R01 NS059690/NS/NINDS NIH HHS/United States
- R01 AG034228/AG/NIA NIH HHS/United States
- F32 NS064596/NS/NINDS NIH HHS/United States
- R01NS059690/NS/NINDS NIH HHS/United States
- P50 AG025688/AG/NIA NIH HHS/United States
- P50AG025688/AG/NIA NIH HHS/United States
- R01 NS038712/NS/NINDS NIH HHS/United States
- 1F32NS064596/NS/NINDS NIH HHS/United States
- 1K99NS064097/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
