

Direct evidence that RNA inhibits APOBEC3G ssDNA cytidine deaminase activity
- PMID: 21856286
- PMCID: PMC3171569
- DOI: 10.1016/j.bbrc.2011.08.009
Direct evidence that RNA inhibits APOBEC3G ssDNA cytidine deaminase activity
Abstract
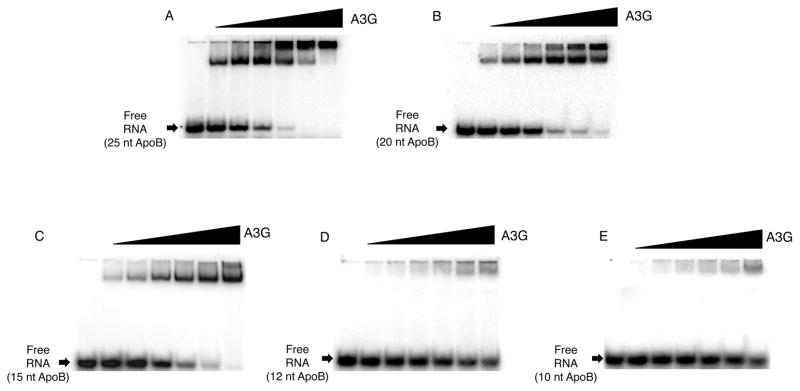
APOBEC3G (A3G) is a deoxycytidine deaminase active on ssDNA substrates. In HIV infected cells A3G interacted with reverse transcription complexes where its activity as a deoxycytidine deaminase led to mutation of the viral genome. A3G not only bound ssDNA, but it also had an intrinsic ability to bind RNA. In many cell types that can support HIV replication, A3G ssDNA deaminase activity was suppressed and the enzyme resided in high molecular mass, ribonucleoprotein complexes associated with cytoplasmic P-bodies and stress granules. Using a defined in vitro system, we show that RNA alone was sufficient to suppress A3G deaminase activity and did so in an RNA concentration-dependent manner. RNAs of diverse sequences and as short as 25nt were effective inhibitors. Native PAGE analyses showed that RNA formed ribonucleoprotein complexes with A3G and in so doing prevented ssDNA substrates from binding to A3G. The data provided direct evidence that A3G binding to cellular RNAs constituted a substantial impediment to the enzyme's ability to interact with ssDNA.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures



References
-
- Harris RS, Petersen-Mahrt SK, Neuberger MS. RNA editing enzyme APOBEC1 and some of its homologs can act as DNA mutators. Mol Cell. 2002;10:1247–1253. - PubMed
-
- Jarmuz A, Chester A, Bayliss J, Gisbourne J, Dunham I, Scott J, Navaratnam N. An Anthropoid-Specific Locus of Orphan C to U RNA-Editing Enzymes on Chromosome 22. Genomics. 2002;79:285–296. - PubMed
-
- Wedekind JE, Dance GS, Sowden MP, Smith HC. Messenger RNA editing in mammals: new members of the APOBEC family seeking roles in the family business. Trends in genetics : TIG. 2003;19:207–216. - PubMed
-
- Newman EN, Holmes RK, Craig HM, Klein KC, Lingappa JR, Malim MH, Sheehy AM. Antiviral function of APOBEC3G can be dissociated from cytidine deaminase activity. Curr Biol. 2005;15:166–170. - PubMed
-
- Hache G, Liddament MT, Harris RS. The retroviral hypermutation specificity of APOBEC3F and APOBEC3G is governed by the C-terminal DNA cytosine deaminase domain. Journal of Biological Chemistry. 2005;280:10920–10924. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
