

Alignment of sarcoplasmic reticulum-mitochondrial junctions with mitochondrial contact points
- PMID: 21856920
- PMCID: PMC3774555
- DOI: 10.1152/ajpheart.00397.2011
Alignment of sarcoplasmic reticulum-mitochondrial junctions with mitochondrial contact points
Abstract
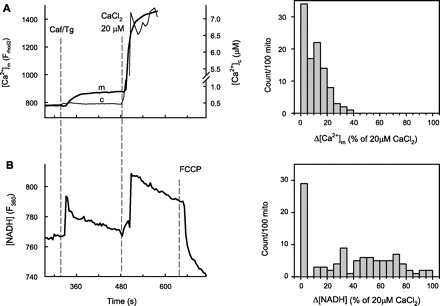
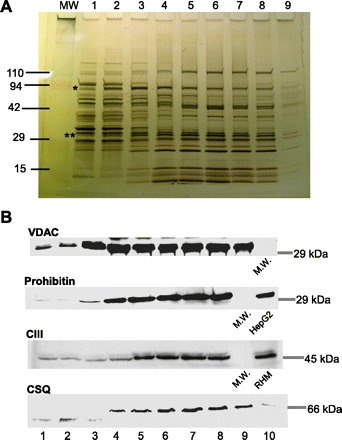
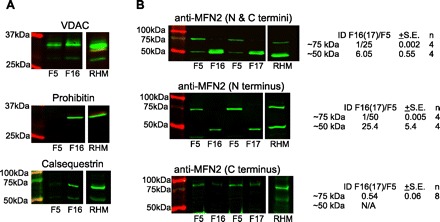
Propagation of ryanodine receptor (RyR2)-derived Ca(2+) signals to the mitochondrial matrix supports oxidative ATP production or facilitates mitochondrial apoptosis in cardiac muscle. Ca(2+) transfer likely occurs locally at focal associations of the sarcoplasmic reticulum (SR) and mitochondria, which are secured by tethers. The outer mitochondrial membrane and inner mitochondrial membrane (OMM and IMM, respectively) also form tight focal contacts (contact points) that are enriched in voltage-dependent anion channels, the gates of OMM for Ca(2+). Contact points could offer the shortest Ca(2+) transfer route to the matrix; however, their alignment with the SR-OMM associations remains unclear. Here, in rat heart we have studied the distribution of mitochondria-associated SR in submitochondrial membrane fractions and evaluated the colocalization of SR-OMM associations with contact points using transmission electron microscopy. In a sucrose gradient designed for OMM purification, biochemical assays revealed lighter fractions enriched in OMM only and heavier fractions containing OMM, IMM, and SR markers. Pure OMM fractions were enriched in mitofusin 2, an ~80 kDa mitochondrial fusion protein and SR-mitochondrial tether candidate, whereas in fractions of OMM + IMM + SR, a lighter (~50 kDa) band detected by antibodies raised against the NH(2) terminus of mitofusin 2 was dominating. Transmission electron microscopy revealed mandatory presence of contact points at the junctional SR-mitochondrial interface versus a random presence along matching SR-free OMM segments. For each SR-mitochondrial junction at least one tether was attached to contact points. These data establish the contact points as anchorage sites for the SR-mitochondrial physical coupling. Close coupling of the SR, OMM, and IMM is likely to provide a favorable spatial arrangement for local ryanodine receptor-mitochondrial Ca(2+) signaling.
Figures




References
-
- Adams V, Bosch W, Schlegel J, Wallimann T, Brdiczka D. Further characterization of contact sites from mitochondria of different tissues: topology of peripheral kinases. Biochim Biophys Acta 981: 213–225, 1989 - PubMed
-
- Ardail D, Gasnier F, Lerme F, Simonot C, Louisot P, Gateau-Roesch O. Involvement of mitochondrial contact sites in the subcellular compartmentalization of phospholipid biosynthetic enzymes. J Biol Chem 268: 25985–25992, 1993 - PubMed
-
- Ardail D, Privat JP, Egret-Charlier M, Levrat C, Lerme F, Louisot P. Mitochondrial contact sites. Lipid composition and dynamics. J Biol Chem 265: 18797–18802, 1990 - PubMed
-
- Bathori G, Csordas G, Garcia-Perez C, Davies E, Hajnoczky G. Ca2+-dependent control of the permeability properties of the mitochondrial outer membrane and voltage-dependent anion-selective channel (VDAC). J Biol Chem 281: 17347–17358, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous