

Increased muscle stress-sensitivity induced by selenoprotein N inactivation in mouse: a mammalian model for SEPN1-related myopathy
- PMID: 21858002
- PMCID: PMC3152547
- DOI: 10.1371/journal.pone.0023094
Increased muscle stress-sensitivity induced by selenoprotein N inactivation in mouse: a mammalian model for SEPN1-related myopathy
Abstract
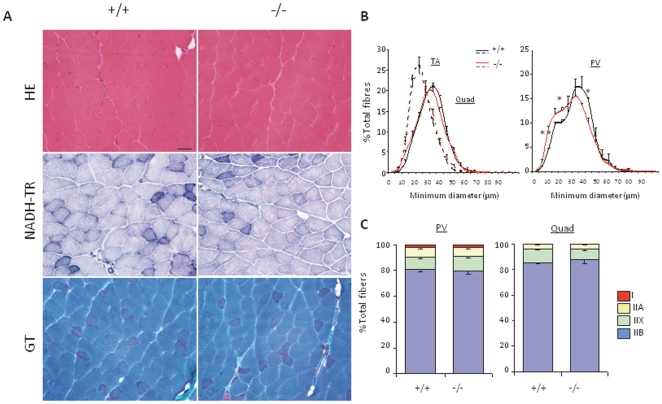
Selenium is an essential trace element and selenoprotein N (SelN) was the first selenium-containing protein shown to be directly involved in human inherited diseases. Mutations in the SEPN1 gene, encoding SelN, cause a group of muscular disorders characterized by predominant affection of axial muscles. SelN has been shown to participate in calcium and redox homeostasis, but its pathophysiological role in skeletal muscle remains largely unknown. To address SelN function in vivo, we generated a Sepn1-null mouse model by gene targeting. The Sepn1(-/-) mice had normal growth and lifespan, and were macroscopically indistinguishable from wild-type littermates. Only minor defects were observed in muscle morphology and contractile properties in SelN-deficient mice in basal conditions. However, when subjected to challenging physical exercise and stress conditions (forced swimming test), Sepn1(-/-) mice developed an obvious phenotype, characterized by limited motility and body rigidity during the swimming session, as well as a progressive curvature of the spine and predominant alteration of paravertebral muscles. This induced phenotype recapitulates the distribution of muscle involvement in patients with SEPN1-Related Myopathy, hence positioning this new animal model as a valuable tool to dissect the role of SelN in muscle function and to characterize the pathophysiological process.
Conflict of interest statement
Figures




References
-
- Rayman MP. Selenoproteins and human health: insights from epidemiological data. Biochim Biophys Acta. 2009;1790:1533–1540. - PubMed
-
- Allmang C, Wurth L, Krol A. The selenium to selenoprotein pathway in eukaryotes: more molecular partners than anticipated. Biochim Biophys Acta. 2009;1790:1415–1423. - PubMed
-
- Arner ES. Selenoproteins-What unique properties can arise with selenocysteine in place of cysteine? Exp Cell Res. 2010;316:1296–1303. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
