

A gain-of-function germline mutation in Drosophila ras1 affects apoptosis and cell fate during development
- PMID: 21858158
- PMCID: PMC3155559
- DOI: 10.1371/journal.pone.0023535
A gain-of-function germline mutation in Drosophila ras1 affects apoptosis and cell fate during development
Abstract
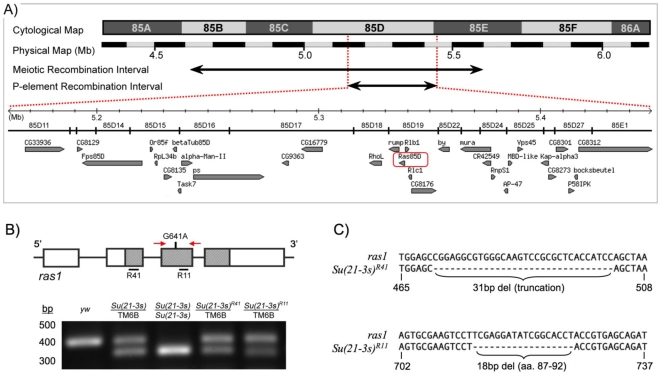
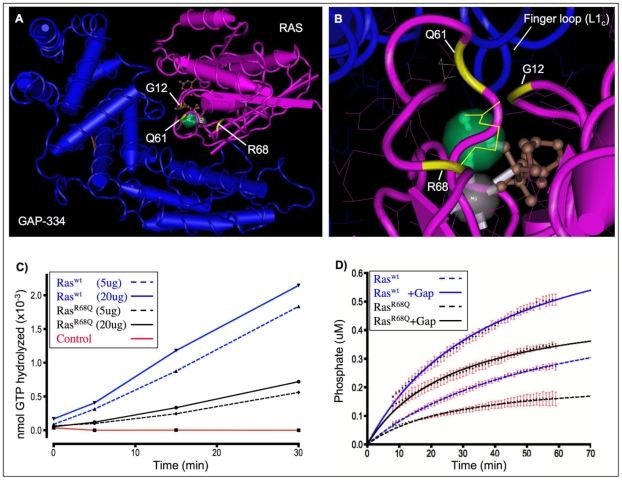
The RAS/MAPK signal transduction pathway is an intracellular signaling cascade that transmits environmental signals from activated receptor tyrosine kinases (RTKs) on the cell surface and other endomembranes to transcription factors in the nucleus, thereby linking extracellular stimuli to changes in gene expression. Largely as a consequence of its role in oncogenesis, RAS signaling has been the subject of intense research efforts for many years. More recently, it has been shown that milder perturbations in Ras signaling during embryogenesis also contribute to the etiology of a group of human diseases. Here we report the identification and characterization of the first gain-of-function germline mutation in Drosophila ras1 (ras85D), the Drosophila homolog of human K-ras, N-ras and H-ras. A single amino acid substitution (R68Q) in the highly conserved switch II region of Ras causes a defective protein with reduced intrinsic GTPase activity, but with normal sensitivity to GAP stimulation. The ras1(R68Q) mutant is homozygous viable but causes various developmental defects associated with elevated Ras signaling, including cell fate changes and ectopic survival of cells in the nervous system. These biochemical and functional properties are reminiscent of germline Ras mutants found in patients afflicted with Noonan, Costello or cardio-facio-cutaneous syndromes. Finally, we used ras1(R68Q) to identify novel genes that interact with Ras and suppress cell death.
Conflict of interest statement
Figures



Similar articles
-
Ras2, the TC21/R-Ras2 Drosophila homologue, contributes to insulin signalling but is not required for organism viability.Dev Biol. 2020 May 15;461(2):172-183. doi: 10.1016/j.ydbio.2020.02.009. Epub 2020 Feb 14. Dev Biol. 2020. PMID: 32061885
-
Cytosolic Ras supports eye development in Drosophila.Mol Cell Biol. 2010 Dec;30(24):5649-57. doi: 10.1128/MCB.00635-10. Epub 2010 Oct 11. Mol Cell Biol. 2010. PMID: 20937772 Free PMC article.
-
A misexpression screen identifies genes that can modulate RAS1 pathway signaling in Drosophila melanogaster.Genetics. 2000 Nov;156(3):1219-30. doi: 10.1093/genetics/156.3.1219. Genetics. 2000. PMID: 11063696 Free PMC article.
-
Deregulated Ras signaling in developmental disorders: new tricks for an old dog.Curr Opin Genet Dev. 2007 Feb;17(1):15-22. doi: 10.1016/j.gde.2006.12.004. Curr Opin Genet Dev. 2007. PMID: 17208427 Review.
-
Noonan, Costello and cardio-facio-cutaneous syndromes: dysregulation of the Ras-MAPK pathway.Expert Rev Mol Med. 2008 Dec 9;10:e37. doi: 10.1017/S1462399408000902. Expert Rev Mol Med. 2008. PMID: 19063751 Review.
Cited by
-
A genome-wide Drosophila epithelial tumorigenesis screen identifies Tetraspanin 29Fb as an evolutionarily conserved suppressor of Ras-driven cancer.PLoS Genet. 2018 Oct 16;14(10):e1007688. doi: 10.1371/journal.pgen.1007688. eCollection 2018 Oct. PLoS Genet. 2018. PMID: 30325918 Free PMC article.
-
Expansion of Drosophila haemocytes using a conditional GeneSwitch driver affects larval haemocyte function, but does not modulate adult lifespan or survival after severe infection.J Exp Biol. 2025 May 1;228(9):jeb249649. doi: 10.1242/jeb.249649. Epub 2025 May 6. J Exp Biol. 2025. PMID: 40116111 Free PMC article.
-
RASopathies: unraveling mechanisms with animal models.Dis Model Mech. 2015 Aug 1;8(8):769-82. doi: 10.1242/dmm.020339. Dis Model Mech. 2015. PMID: 26203125 Free PMC article. Review.
-
Habituation Learning Is a Widely Affected Mechanism in Drosophila Models of Intellectual Disability and Autism Spectrum Disorders.Biol Psychiatry. 2019 Aug 15;86(4):294-305. doi: 10.1016/j.biopsych.2019.04.029. Epub 2019 May 9. Biol Psychiatry. 2019. PMID: 31272685 Free PMC article.
References
-
- Cantley LC, Auger KR, Carpenter C, Duckworth B, Graziani A, et al. Oncogenes and signal transduction. Cell. 1991;64:281–302. - PubMed
-
- Blume-Jensen P, Hunter T. Oncogenic kinase signalling. Nature. 2001;411:355–365. - PubMed
-
- Bishop JM. Molecular themes in oncogenesis. Cell. 1991;64:235–248. - PubMed
-
- Hunter T. Cooperation between oncogenes. Cell. 1991;64:249–270. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
