

Role of reuniens nucleus projections to the medial prefrontal cortex and to the hippocampal pyramidal CA1 area in associative learning
- PMID: 21858159
- PMCID: PMC3156136
- DOI: 10.1371/journal.pone.0023538
Role of reuniens nucleus projections to the medial prefrontal cortex and to the hippocampal pyramidal CA1 area in associative learning
Abstract
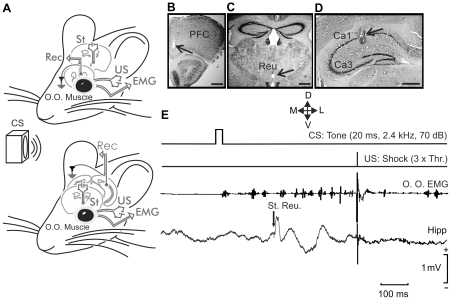
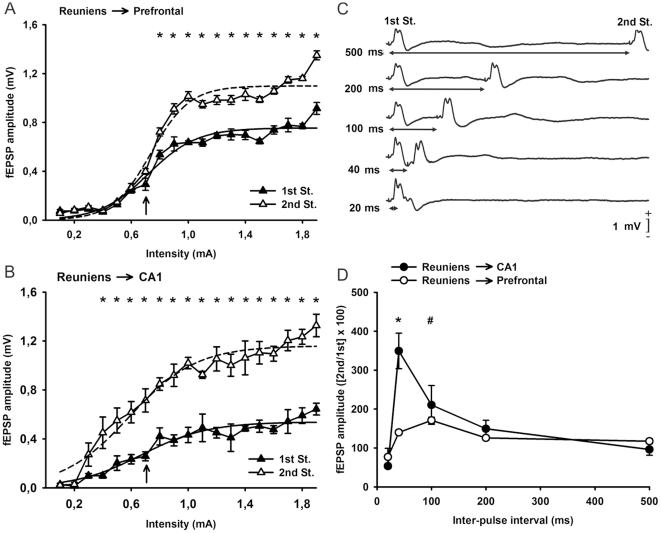
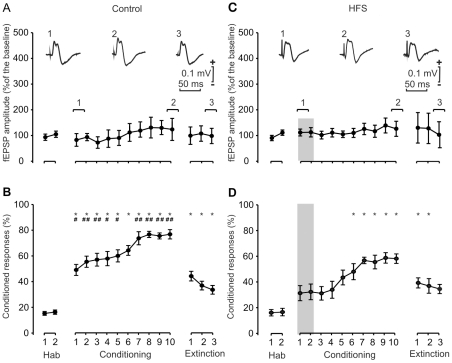
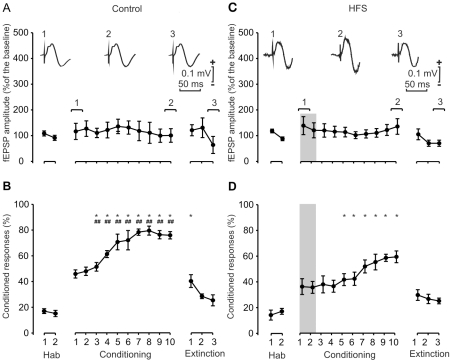
We studied the interactions between short- and long-term plastic changes taking place during the acquisition of a classical eyeblink conditioning and following high-frequency stimulation (HFS) of the reuniens nucleus in behaving mice. Synaptic changes in strength were studied at the reuniens-medial prefrontal cortex (mPFC) and the reuniens-CA1 synapses. Input/output curves and a paired-pulse study enabled determining the functional capabilities of the two synapses and the optimal intensities to be applied at the reuniens nucleus during classical eyeblink conditioning and for HFS applied to the reuniens nucleus. Animals were conditioned using a trace paradigm, with a tone as conditioned stimulus (CS) and an electric shock to the trigeminal nerve as unconditioned stimulus (US). A single pulse was presented to the reuniens nucleus to evoke field EPSPs (fEPSPs) in mPFC and CA1 areas during the CS-US interval. No significant changes in synaptic strength were observed at the reuniens-mPFC and reuniens-CA1 synapses during the acquisition of eyelid conditioned responses (CRs). Two successive HFS sessions carried out during the first two conditioning days decreased the percentage of CRs, without evoking any long-term potentiation (LTP) at the recording sites. HFS of the reuniens nucleus also prevented the proper acquisition of an object discrimination task. A subsequent study revealed that HFS of the reuniens nucleus evoked a significant decrease of paired-pulse facilitation. In conclusion, reuniens nucleus projections to prefrontal and hippocampal circuits seem to participate in the acquisition of associative learning through a mechanism that does not required the development of LTP.
Conflict of interest statement
Figures


Similar articles
-
Involvement of the CA3-CA1 synapse in the acquisition of associative learning in behaving mice.J Neurosci. 2006 Jan 25;26(4):1077-87. doi: 10.1523/JNEUROSCI.2834-05.2006. J Neurosci. 2006. PMID: 16436593 Free PMC article.
-
Differential effects of long-term potentiation evoked at the CA3 CA1 synapse before, during, and after the acquisition of classical eyeblink conditioning in behaving mice.J Neurosci. 2007 Nov 7;27(45):12139-46. doi: 10.1523/JNEUROSCI.3397-07.2007. J Neurosci. 2007. PMID: 17989280 Free PMC article.
-
Adenosine A(2A) receptor modulation of hippocampal CA3-CA1 synapse plasticity during associative learning in behaving mice.Neuropsychopharmacology. 2009 Jun;34(7):1865-74. doi: 10.1038/npp.2009.8. Epub 2009 Feb 11. Neuropsychopharmacology. 2009. PMID: 19212319
-
Functional basis of associative learning and its relationships with long-term potentiation evoked in the involved neural circuits: Lessons from studies in behaving mammals.Neurobiol Learn Mem. 2015 Oct;124:3-18. doi: 10.1016/j.nlm.2015.04.006. Epub 2015 Apr 25. Neurobiol Learn Mem. 2015. PMID: 25916668 Review.
-
Activity-dependent changes of the hippocampal CA3-CA1 synapse during the acquisition of associative learning in conscious mice.Genes Brain Behav. 2007 Jun;6 Suppl 1:24-31. doi: 10.1111/j.1601-183X.2007.00319.x. Genes Brain Behav. 2007. PMID: 17543036 Review.
Cited by
-
Prefrontal-Hippocampal Interactions in Memory and Emotion.Front Syst Neurosci. 2015 Dec 15;9:170. doi: 10.3389/fnsys.2015.00170. eCollection 2015. Front Syst Neurosci. 2015. PMID: 26696844 Free PMC article. Review.
-
Nucleus reuniens of the thalamus contains head direction cells.Elife. 2014 Jul 14;3:e03075. doi: 10.7554/eLife.03075. Elife. 2014. PMID: 25024427 Free PMC article.
-
Nucleus reuniens transiently synchronizes memory networks at beta frequencies.Nat Commun. 2023 Jul 19;14(1):4326. doi: 10.1038/s41467-023-40044-z. Nat Commun. 2023. PMID: 37468487 Free PMC article.
-
A Critical Role for the Nucleus Reuniens in Long-Term, But Not Short-Term Associative Recognition Memory Formation.J Neurosci. 2018 Mar 28;38(13):3208-3217. doi: 10.1523/JNEUROSCI.1802-17.2017. Epub 2018 Feb 15. J Neurosci. 2018. PMID: 29449430 Free PMC article.
-
The reuniens and rhomboid nuclei: neuroanatomy, electrophysiological characteristics and behavioral implications.Prog Neurobiol. 2013 Dec;111:34-52. doi: 10.1016/j.pneurobio.2013.08.006. Epub 2013 Sep 8. Prog Neurobiol. 2013. PMID: 24025745 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
