

Regulation of anthrax toxin-specific antibody titers by natural killer T cell-derived IL-4 and IFNγ
- PMID: 21858226
- PMCID: PMC3157475
- DOI: 10.1371/journal.pone.0023817
Regulation of anthrax toxin-specific antibody titers by natural killer T cell-derived IL-4 and IFNγ
Abstract
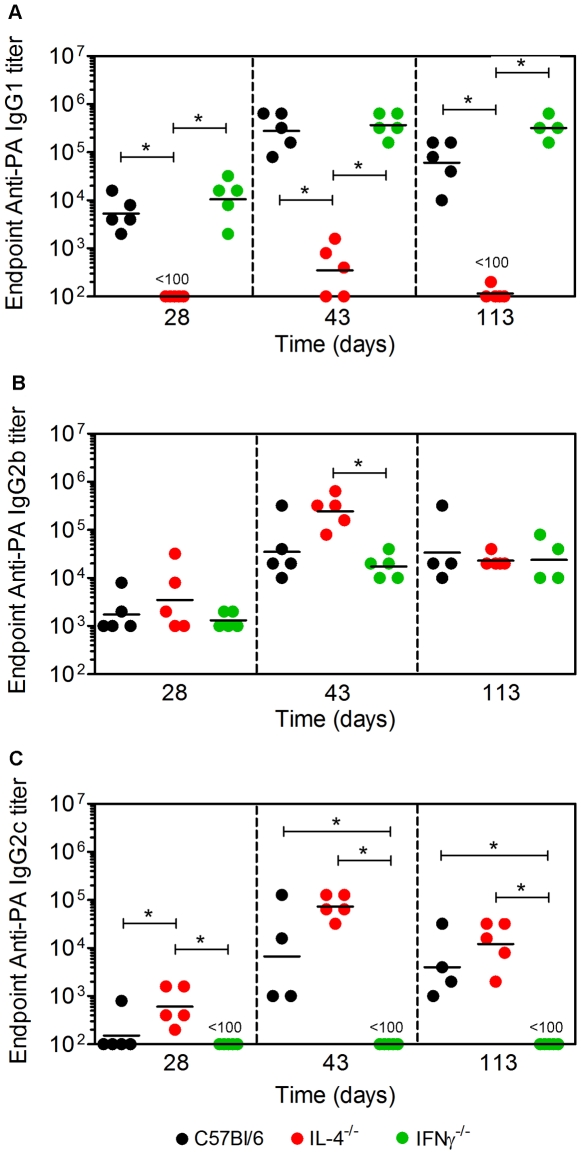
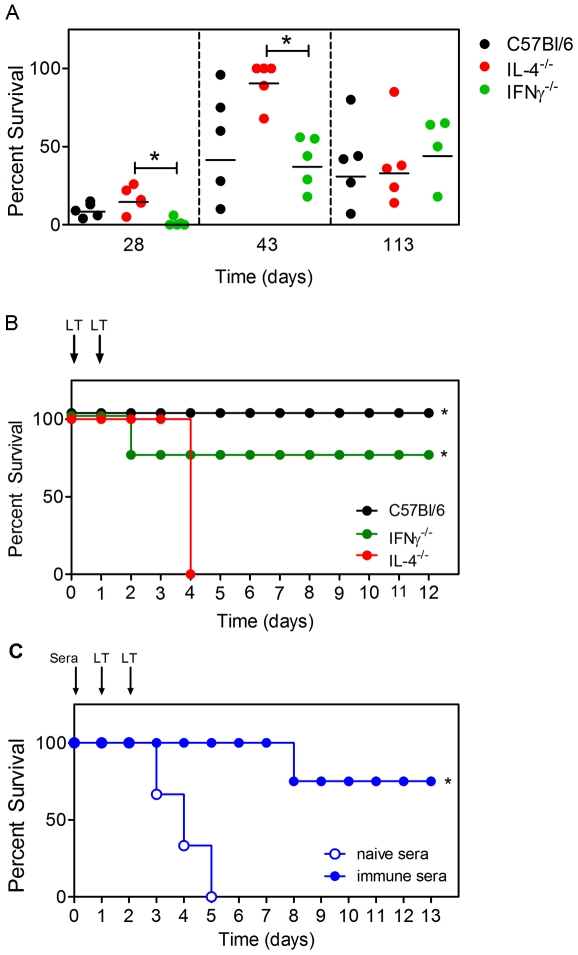
Activation of Natural Killer-like T cells (NKT) with the CD1d ligand α-GC leads to enhanced production of anthrax toxin protective Ag (PA)-neutralizing Abs, yet the underlying mechanism for this adjuvant effect is not known. In the current study we examined the role of Th1 and Th2 type responses in NKT-mediated enhancement of antibody responses to PA. First, the contribution of IL-4 and IFNγ to the production of PA-specific toxin-neutralizing Abs was examined. By immunizing C57Bl/6 controls IL-4(-/-) mice and IFNγ(-/-) mice and performing passive serum transfer experiments, it was observed that sera containing PA-specific IgG1, IgG2b and IgG2c neutralized toxin in vitro and conferred protection in vivo. Sera containing IgG2b and IgG2c neutralized toxin in vitro but were not sufficient for protection in vivo. Sera containing IgG1 and IgG2b neutralized toxin in vitro and conferred protection in vivo. IgG1 therefore emerged as a good correlate of protection. Next, C57Bl/6 mice were immunized with PA alone or PA plus a Th2-skewing α-GC derivative known as OCH. Neutralizing PA-specific IgG1 responses were modestly enhanced by OCH in C57Bl/6 mice. Conversely, IgG2b and IgG2c were considerably enhanced in PA/OCH-immunized IL-4(-/-) mice but did not confer protection. Finally, bone marrow chimeras were generated such that NKT cells were unable to express IL-4 or IFNγ. NKT-derived IL-4 was required for OCH-enhanced primary IgG1 responses but not recall responses. NKT-derived IL-4 and IFNγ also influenced primary and recall IgG2b and IgG2c titers. These data suggest targeted skewing of the Th2 response by α-GC derivatives can be exploited to optimize anthrax vaccination.
Conflict of interest statement
Figures
References
-
- Bradley KA, Mogridge J, Mourez M, Collier RJ, Young JA. Identification of the cellular receptor for anthrax toxin. Nature. 2001;414:225–229. - PubMed
-
- Collier RJ, Young JA. Anthrax toxin. Annu Rev Cell Dev Biol. 2003;19:45–70. - PubMed
-
- Leppla SH. Bacillus anthracis calmodulin-dependent adenylate cyclase: chemical and enzymatic properties and interactions with eucaryotic cells. Adv Cyclic Nucleotide Protein Phosphorylation Res. 1984;17:189–198. - PubMed
-
- Mock M, Mignot T. Anthrax toxins and the host: a story of intimacy. Cell Microbiol. 2003;5:15–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
