

Review
doi: 10.1038/nrm3175.
Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis
Affiliations
- PMID: 21860392
- PMCID: PMC3280198
- DOI: 10.1038/nrm3175
Item in Clipboard
Review
Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis
Nat Rev Mol Cell Biol.
.
Abstract
To provide a stable environmental barrier, the epidermis requires an integrated network of cytoskeletal elements and cellular junctions. Nevertheless, the epidermis ranks among the body's most dynamic tissues, continually regenerating itself and responding to cutaneous insults. As keratinocytes journey from the basal compartment towards the cornified layers, they completely reorganize their adhesive junctions and cytoskeleton. These architectural components are more than just rivets and scaffolds - they are active participants in epidermal morphogenesis that regulate epidermal polarization, signalling and barrier formation.
Figures
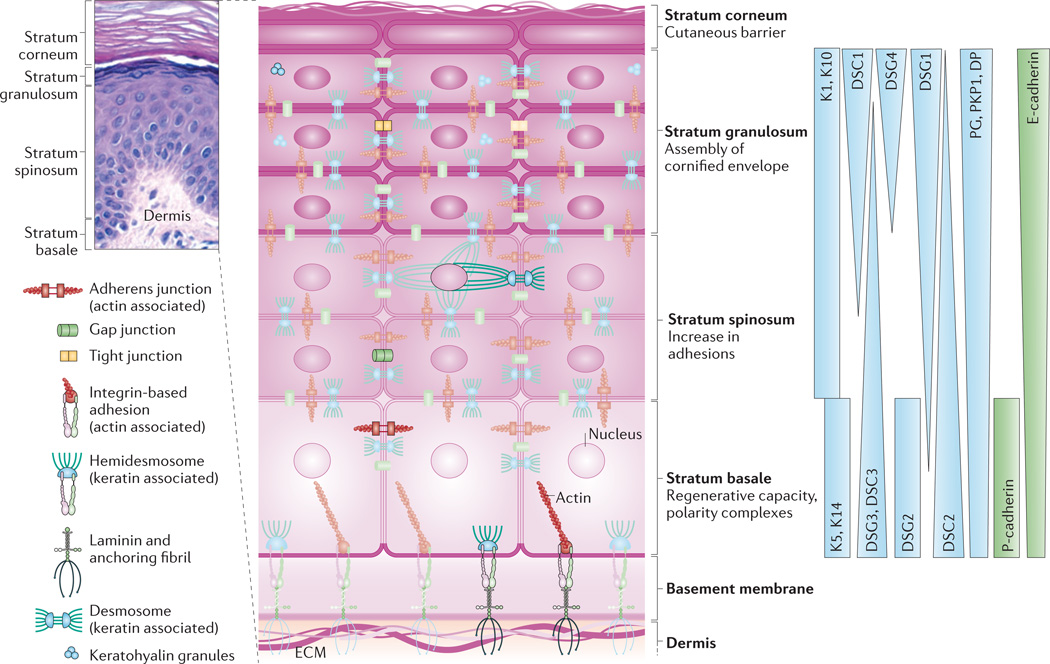
The epidermis is composed of stratified cell layers, which undergo programmed differentiation to allow for constant renewal of the skin. Four main layers are illustrated by a hematoxylin- and eosin-stained human skin sample and an accompanying schematic of the stratum basale, stratum spinosum, stratum granulosum and stratum corneum. The basal, proliferating cell layer of the epidermis remains in contact with the dermis through hemidesmosomes and integrin-based adhesions, both of which provide connections to the underlying extracellular matrix (ECM). During keratinocyte differentiation, a unique cytoarchitecture is elaborated in each of the four layers that comprises specific cytoskeleton and cell junction types, including adherens junctions, tight junctions, desmosomes and gap junctions. The differentiation-dependent changes in the composition and organization of epidermal cytoarchitecture help to drive tissue morphogenesis while supporting the specific functions of each layer, from the regenerative capacity of the stratum basale to the assembly of the cornified envelope and the sloughing of terminally differentiated cells from the stratum corneum. The graded distribution of specific cytoskeletal and junction components, including specific keratins (Ks), desmogleins (DSGs) and cadherins, is crucial for driving morphogenesis. Image in top left courtesy of R. Lavker, Northwestern University, USA. DP, desmoplakin; DSC, desmocollin; E-cadherin, epithelial cadherin; P-cadherin, placental cadherin; PG, plakoglobin; PKP1, plakophilin 1.
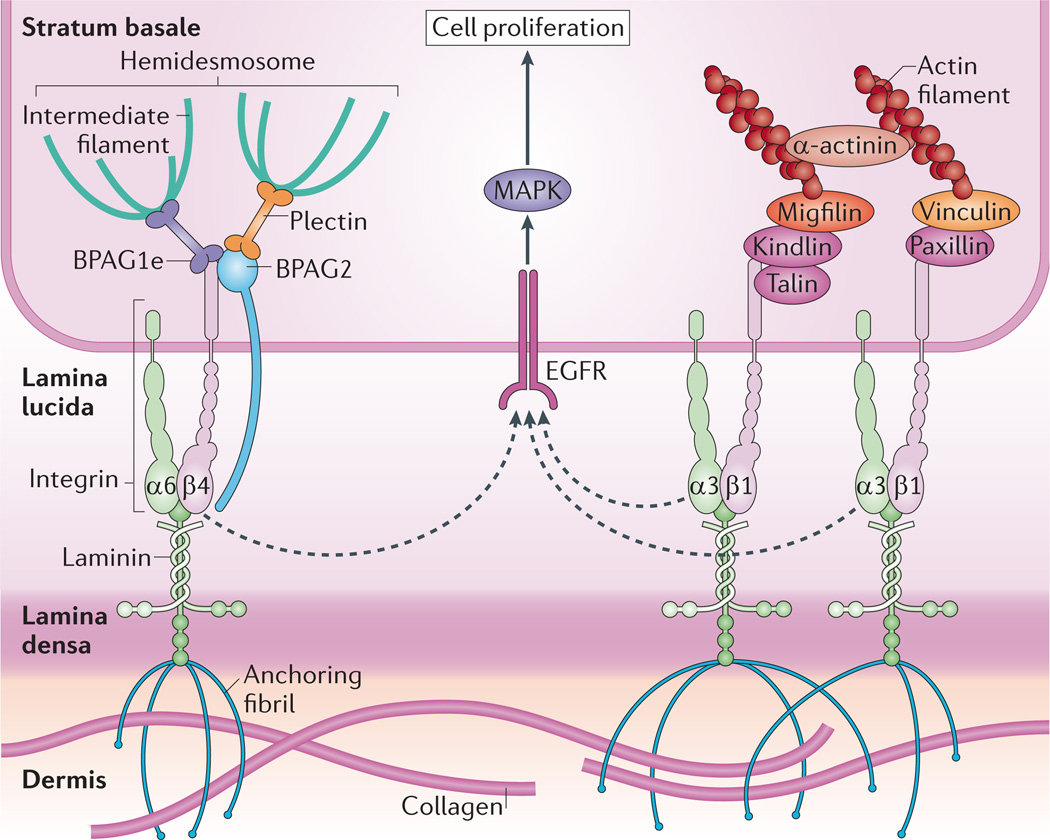
The stratum basale has a high density of hemidesmosomes and integrin-based adhesions that maintain cell attachment to the underlying basement membrane, which is composed of the lamina lucida and lamina densa. Hemidesmosomes connect to the basement membrane through α6β4 integrins and the transmembrane protein bullous pemphigoid antigen 2 (BPAG2; also known as collagen XVII), and are tethered to intermediate filaments by the plakin family members plectin and BPAG1e. α3β1 integrins provide transmembrane connections to the intracellular actin network through recruitment of several factors to their cytoplasmic tails. Integrins are thought to crosstalk with receptors such as EGFR (epidermal growth factor receptor) to induce proliferation of cells in the stratum basale via mitogen-activated protein kinase (MAPK) signalling. As cells stratify, decreased integrin density allows cell cycle exit and differentiation.
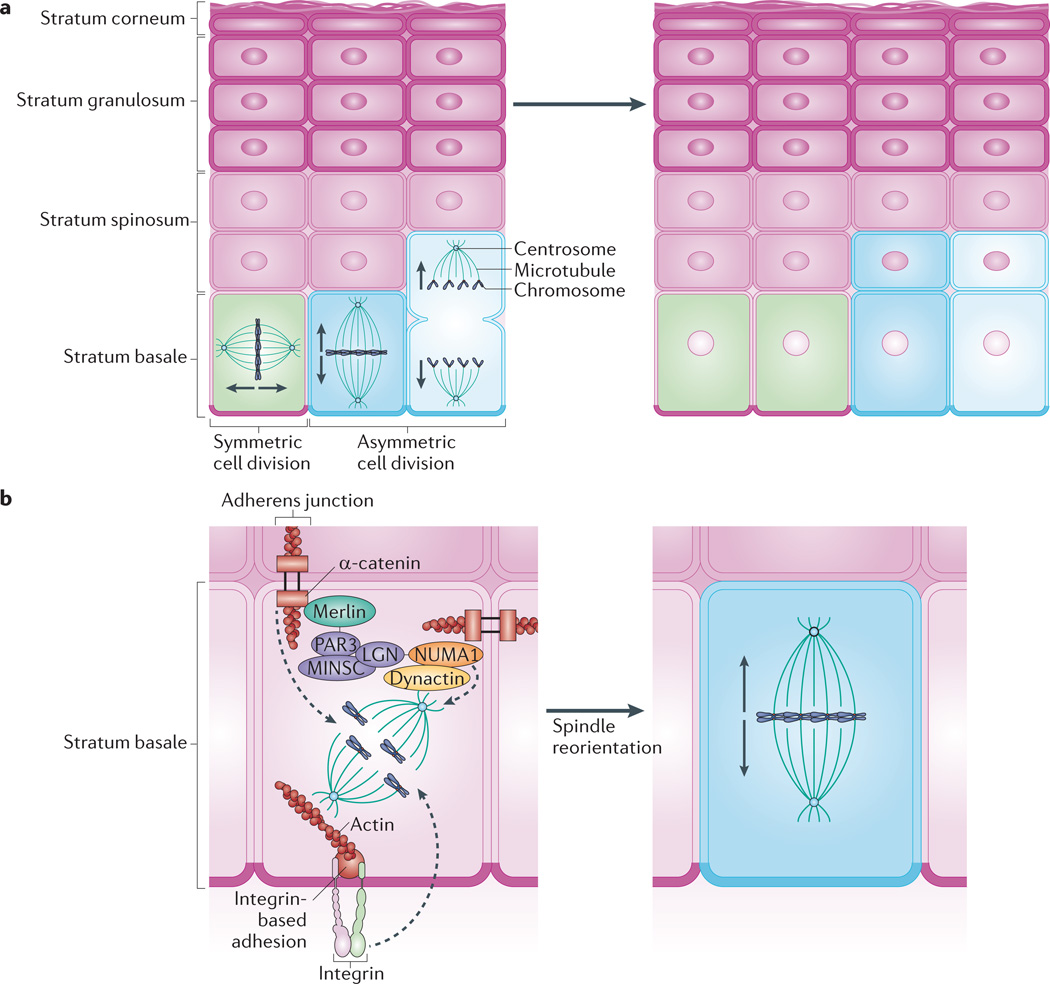
Keratinocytes of the stratum basale provide the regenerative capacity of the epidermis. a | Basal cells use symmetric cell division to promote lateral expansion of the epidermis and undergo asymmetric cell division to enable vertical expansion through the production of differentiated keratinocytes. The direction of expansion is thought to depend on the orientation of the mitotic spindle: spindles that lie parallel to the basement membrane favour symmetric division, whereas spindles perpendicular to the basement membrane promote asymmetric division. b | Asymmetric cell division depends on apical polarity factors, including the partitioning defective 3 (PAR3)–mouse inscuteable (MINSC)–LGN complex, which interacts with nuclear mitotic apparatus protein 1 (NUMA1) and dynactin to regulate spindle orientation,. The adherens junction component α-catenin is thought to recruit this complex via merlin, which links α-catenin and PAR3 (REF. 75). Through knockout studies, p120 catenin and β1 integrin have also been shown to have a role in mitotic spindle orientation,. The roles of these factors in regulating mitotic spindle pole orientation and subsequent division, including the precise location of complexes within basal cells, are yet to be fully elucidated.
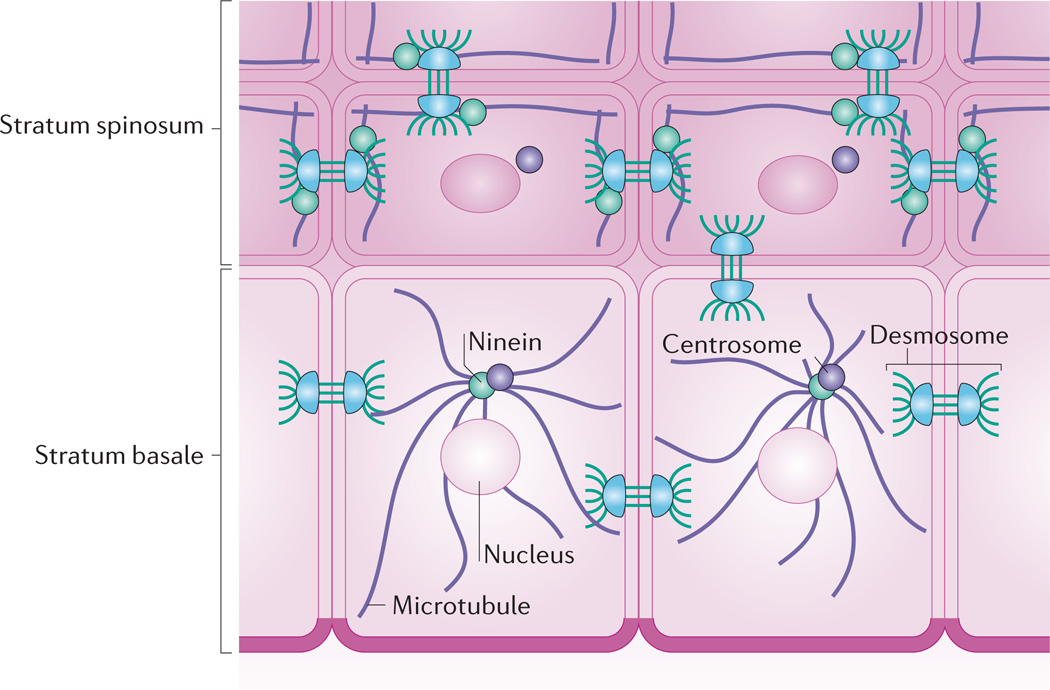
Whereas keratinocytes of the stratum basale have an astral array of microtubules emanating from perinuclear centrosomes, microtubules in suprabasal keratinocytes reorganize to lie parallel with cell borders. This reorganization depends on ninein, a protein that localizes to centrosomes but is recruited to desmosomes in suprabasal keratinocytes through association with desmoplakin.
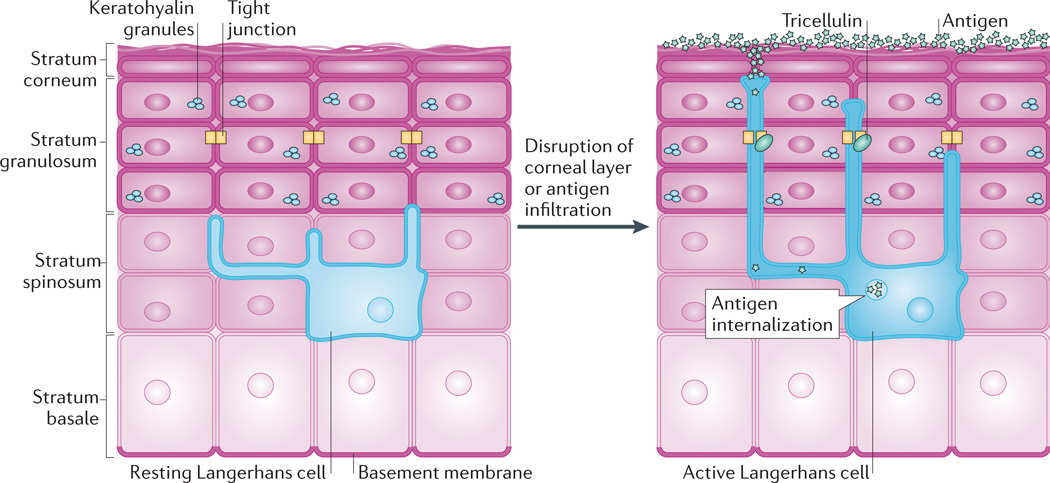
Tight junctions contribute to the barrier between the superficial epidermis and underlying stratum spinosum but must also be dynamic to allow antigen sampling. In the resting state, Langerhans cells reside among keratinocytes in the stratum spinosum and extend dendrites through suprabasal layers. When activated, for instance by disruption of the stratum corneum or by infiltration of antigens, the dendrites of Langerhans cells dock with tight junctions and gain the ability to extend into the stratum corneum. The transmembrane protein tricellulin localizes to these tricellular tight junctions formed between keratinocytes and Langerhans cells. This dynamic adjustment of tight junctions allows Langerhans cells to internalize antigens, which can then be presented to the host immune system.
References
-
- Fuchs E, Raghavan S. Getting under the skin of epidermal morphogenesis. Nature Rev. Genet. 2002;3:199–209. - PubMed
-
- Candi E, Schmidt R, Melino G. The cornified envelope: a model of cell death in the skin. Nature Rev. Mol. Cell Biol. 2005;6:328–340. - PubMed
-
- Mack JA, Anand S, Maytin EV. Proliferation and cornification during development of the mammalian epidermis. Birth Defects Res. C Embryo Today. 2005;75:314–329. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
