

Functions of the poly(ADP-ribose) polymerase superfamily in plants
- PMID: 21861184
- PMCID: PMC11114847
- DOI: 10.1007/s00018-011-0793-4
Functions of the poly(ADP-ribose) polymerase superfamily in plants
Abstract
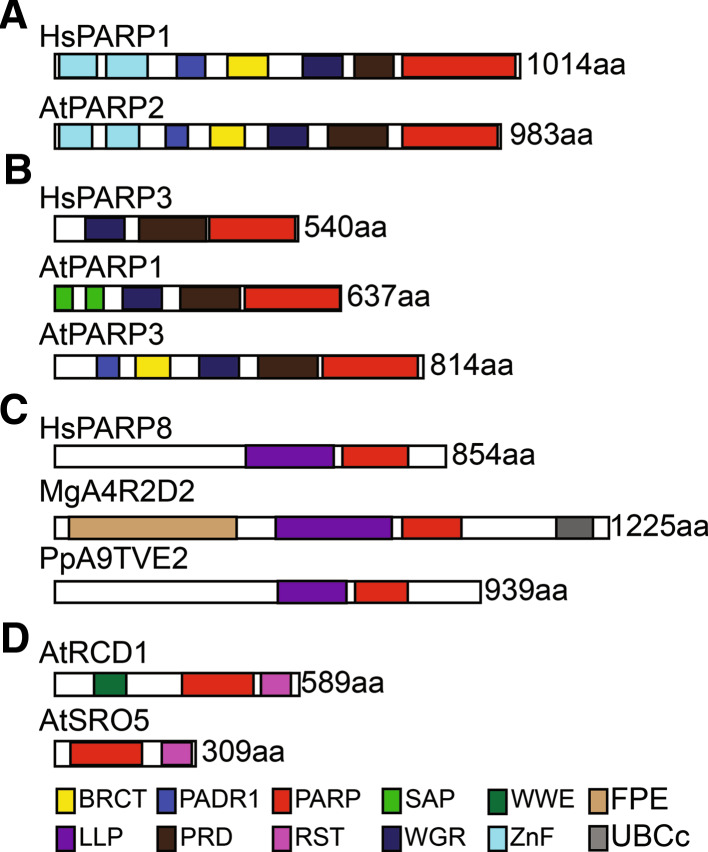
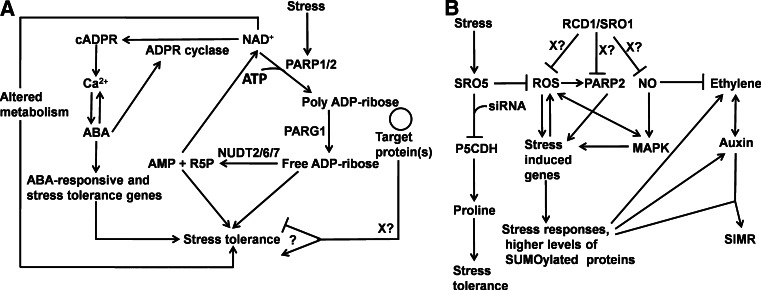
Poly(ADP-ribosyl)ation is the covalent attachment of ADP-ribose subunits from NAD(+) to target proteins and was first described in plants in the 1970s. This post-translational modification is mediated by poly(ADP-ribose) polymerases (PARPs) and removed by poly(ADP-ribose) glycohydrolases (PARGs). PARPs have important functions in many biological processes including DNA repair, epigenetic regulation and transcription. However, these roles are not always associated with enzymatic activity. The PARP superfamily has been well studied in animals, but remains under-investigated in plants. Although plants lack the variety of PARP superfamily members found in mammals, they do encode three different types of PARP superfamily proteins, including a group of PARP-like proteins, the SRO family, that are plant specific. In plants, members of the PARP family and/or poly(ADP-ribosyl)ation have been linked to DNA repair, mitosis, innate immunity and stress responses. In addition, members of the SRO family have been shown to be necessary for normal sporophytic development. In this review, we summarize the current state of plant research into poly(ADP-ribosyl)ation and the PARP superfamily in plants.
Figures
References
-
- Adams-Phillips L, Wan J, Tan X, Dunning FM, Meyers BC, Michelmore RW, Bent AF. Discovery of ADP-ribosylation and other plant defense pathway elements through expression profiling of four different Arabidopsis-Pseudomonas R-avr interactions. Mol Plant Microbe Interact. 2008;21:646–657. doi: 10.1094/MPMI-21-5-0646. - DOI - PubMed
-
- Aguiar RC, Yakushijin Y, Kharbanda S, Salgia R, Fletcher JA, Shipp MA. BAL is a novel risk-related gene in diffuse large B-cell lymphomas that enhances cellular migration. Blood. 2000;96:4328–4334. - PubMed
-
- Ahlfors R, Brosche M, Kollist H, Kangasjarvi J. Nitric oxide modulates ozone-induced cell death, hormone biosynthesis and gene expression in Arabidopsis thaliana. Plant J. 2008;58:1–12. - PubMed
-
- Ahlfors R, Lang S, Overmyer K, Jaspers P, Brosche M, Tauriainen A, Kollist H, Tuominen H, Belles-Boix E, Piippo M, Inze D, Palva ET, Kangasjarvi J. Arabidopsis RADICAL-INDUCED CELL DEATH1 belongs to the WWE protein-protein interaction domain protein family and modulates abscisic acid, ethylene, and methyl jasmonate responses. Plant Cell. 2004;16:1925–1937. doi: 10.1105/tpc.021832. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
