

Elucidation of the pathway to astaxanthin in the flowers of Adonis aestivalis
- PMID: 21862704
- PMCID: PMC3180810
- DOI: 10.1105/tpc.111.086827
Elucidation of the pathway to astaxanthin in the flowers of Adonis aestivalis
Abstract
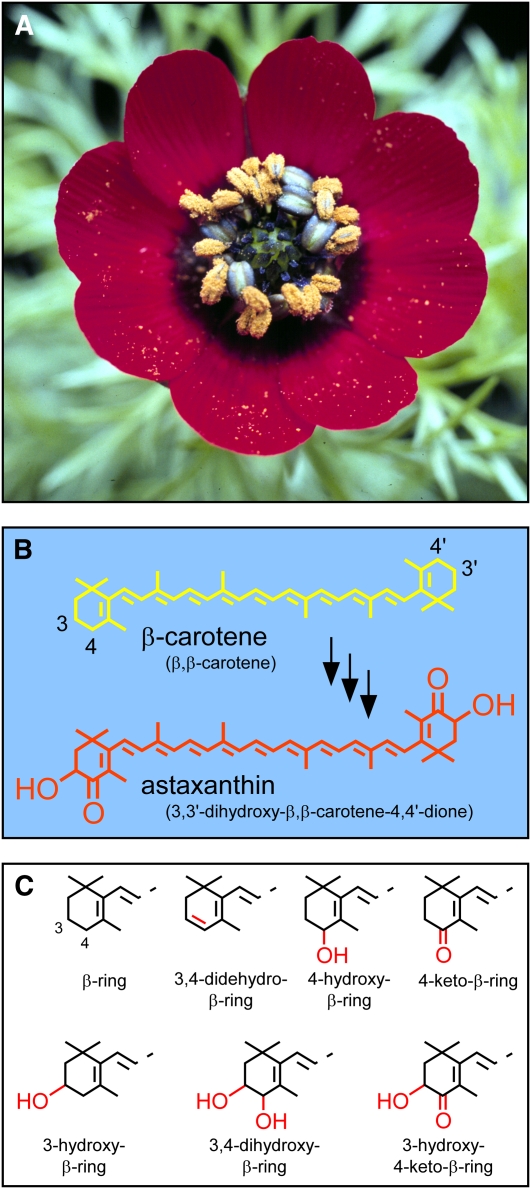
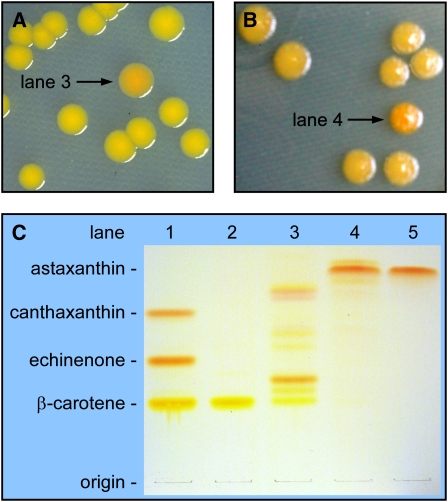
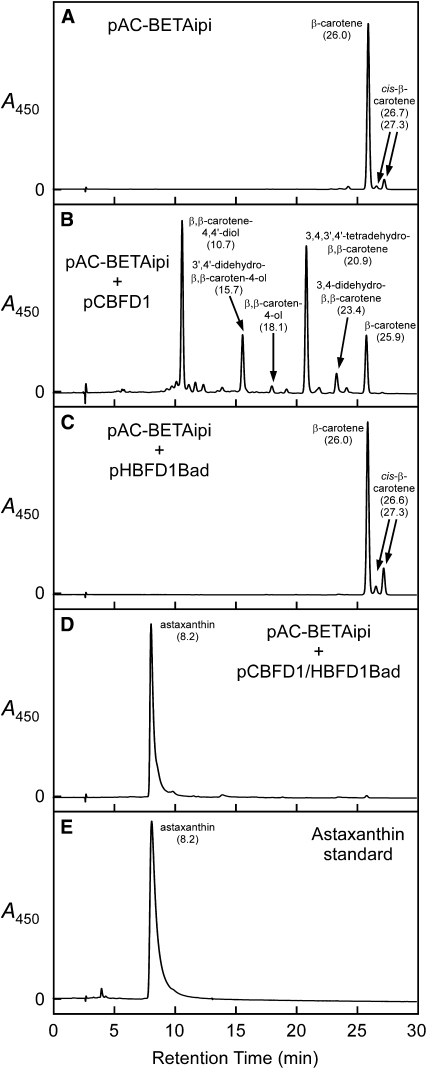
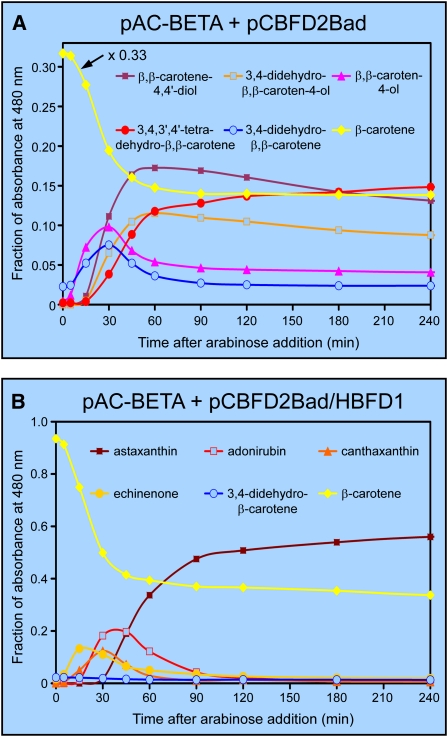
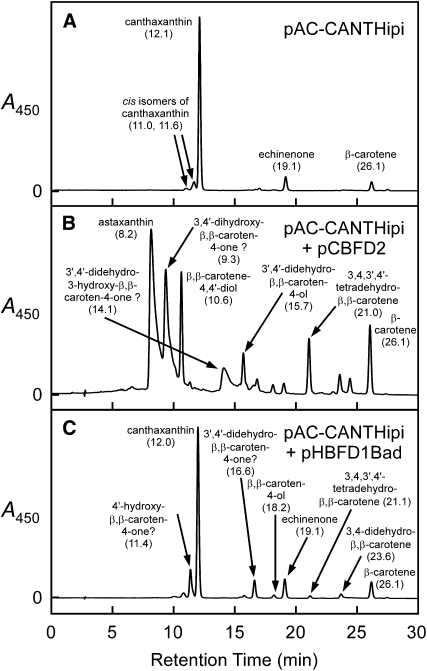
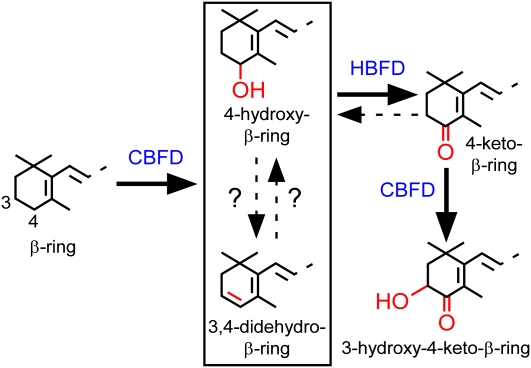
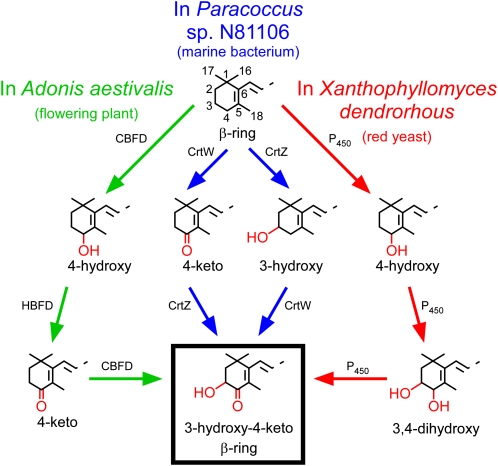
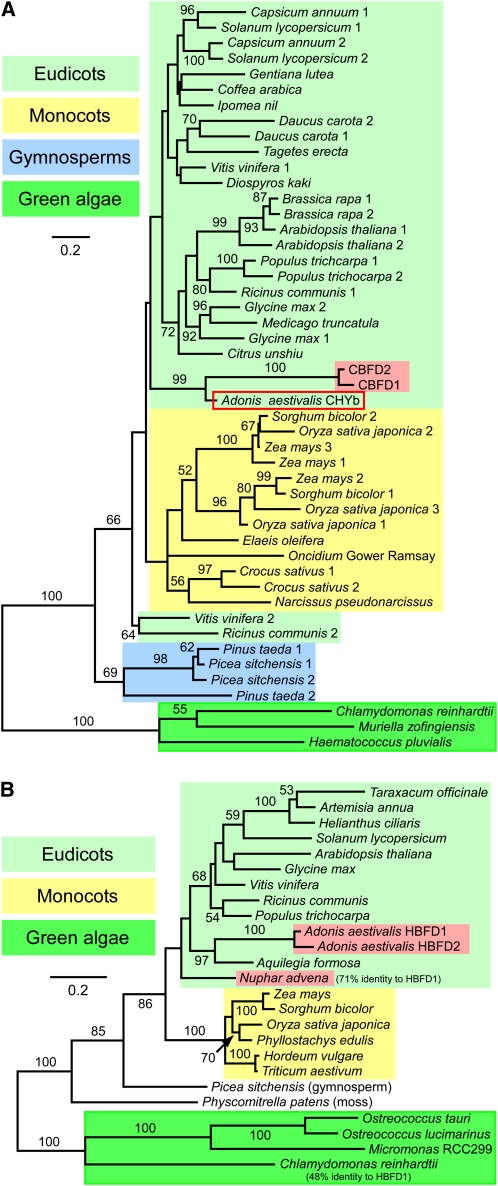
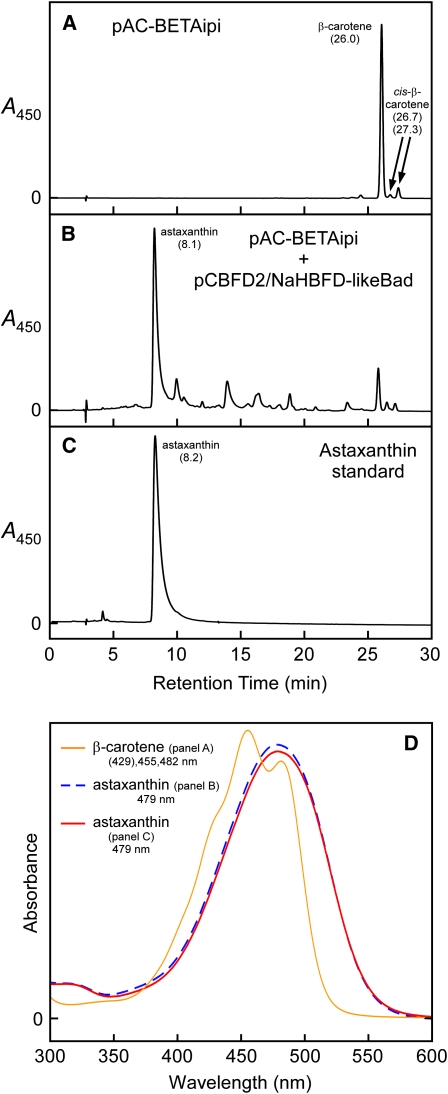
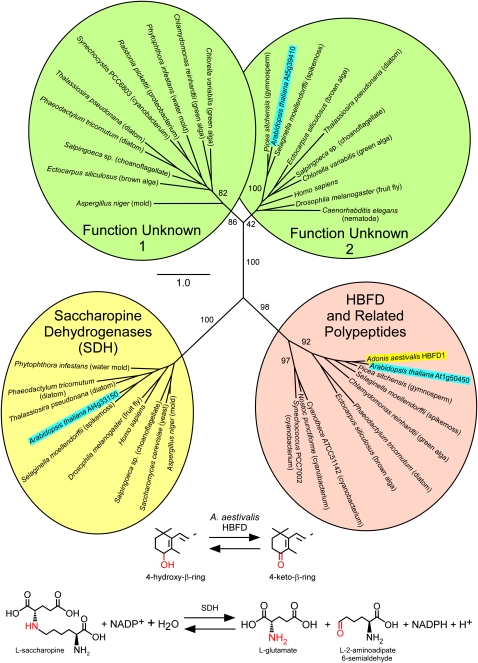
A few species in the genus Adonis are the only land plants known to produce the valuable red ketocarotenoid astaxanthin in abundance. Here, we ascertain the pathway that leads from the β-rings of β-carotene, a carotenoid ubiquitous in plants, to the 3-hydroxy-4-keto-β-rings of astaxanthin (3,3'-dihydroxy-β,β-carotene-4,4'-dione) in the blood-red flowers of Adonis aestivalis, an ornamental and medicinal plant commonly known as summer pheasant's eye. Two gene products were found to catalyze three distinct reactions, with the first and third reactions of the pathway catalyzed by the same enzyme. The pathway commences with the activation of the number 4 carbon of a β-ring in a reaction catalyzed by a carotenoid β-ring 4-dehydrogenase (CBFD), continues with the further dehydrogenation of this carbon to yield a carbonyl in a reaction catalyzed by a carotenoid 4-hydroxy-β-ring 4-dehydrogenase, and concludes with the addition of an hydroxyl group at the number 3 carbon in a reaction catalyzed by the erstwhile CBFD enzyme. The A. aestivalis pathway is both portable and robust, functioning efficiently in a simple bacterial host. Our elucidation of the pathway to astaxanthin in A. aestivalis provides enabling technology for development of a biological production process and reveals the evolutionary origin of this unusual plant pathway, one unrelated to and distinctly different from those used by bacteria, green algae, and fungi to synthesize astaxanthin.
Figures
References
-
- Álvarez V., Rodríguez-Sáiz M., de la Fuente J.L., Gudiña E.J., Godio R.P., Martín J.F., Barredo J.L. (2006). The crtS gene of Xanthophyllomyces dendrorhous encodes a novel cytochrome-P450 hydroxylase involved in the conversion of β-carotene into astaxanthin and other xanthophylls. Fungal Genet. Biol. 43: 261–272 - PubMed
-
- Andi B., Cook P.F., West A.H. (2006). Crystal structure of the his-tagged saccharopine reductase from Saccharomyces cerevisiae at 1.7-Å resolution. Cell Biochem. Biophys. 46: 17–26 - PubMed
-
- Bauernfeind J.C., Adams C.R., Marusich W.L. (1981). Carotenes and other vitamin A precursors in animal feed. In Carotenoids as Colorants and Vitamin A Precursors: Technological and Nutritional Applications, Bauernfeind J.C., (New York: Academic Press; ), pp. 563–743
-
- Broadwater J.A., Whittle E., Shanklin J. (2002). Desaturation and hydroxylation. Residues 148 and 324 of Arabidopsis FAD2, in addition to substrate chain length, exert a major influence in partitioning of catalytic specificity. J. Biol. Chem. 277: 15613–15620 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
