

Generation of signaling specificity in Arabidopsis by spatially restricted buffering of ligand-receptor interactions
- PMID: 21862708
- PMCID: PMC3180797
- DOI: 10.1105/tpc.111.086637
Generation of signaling specificity in Arabidopsis by spatially restricted buffering of ligand-receptor interactions
Abstract
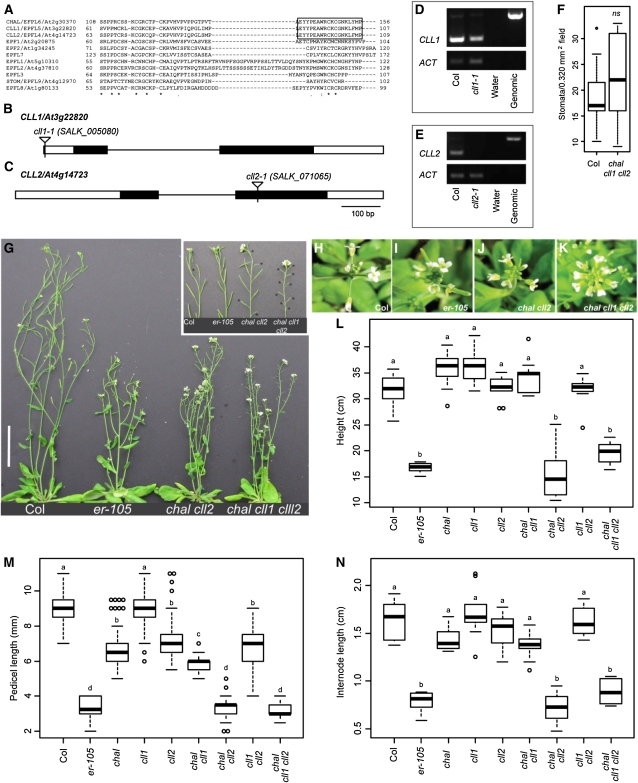
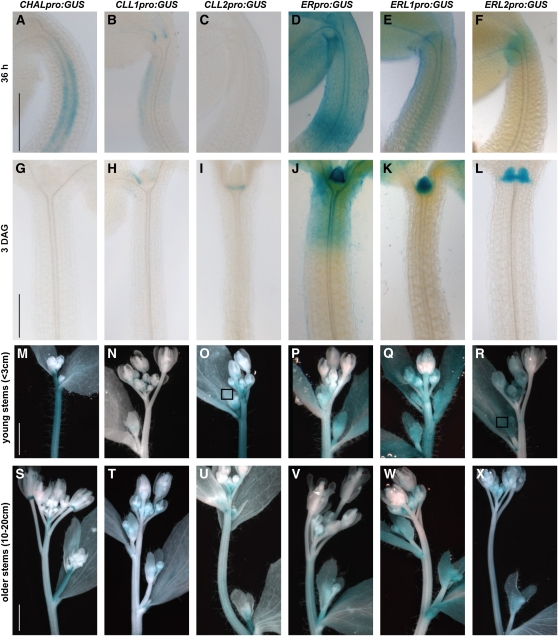
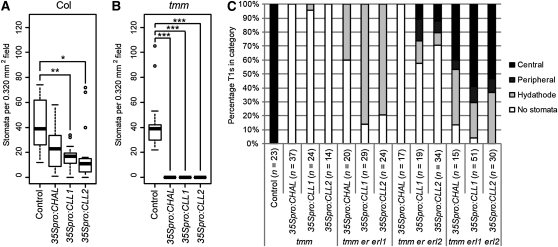
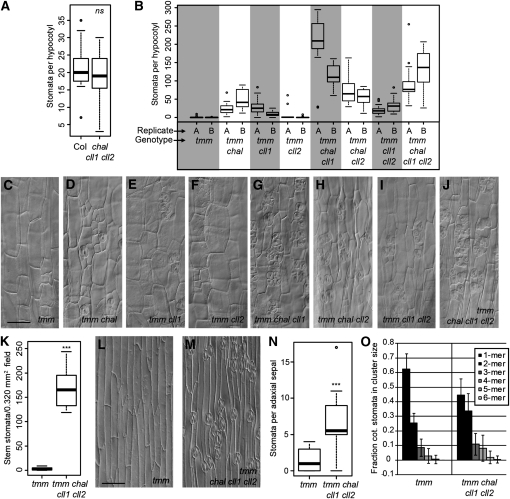
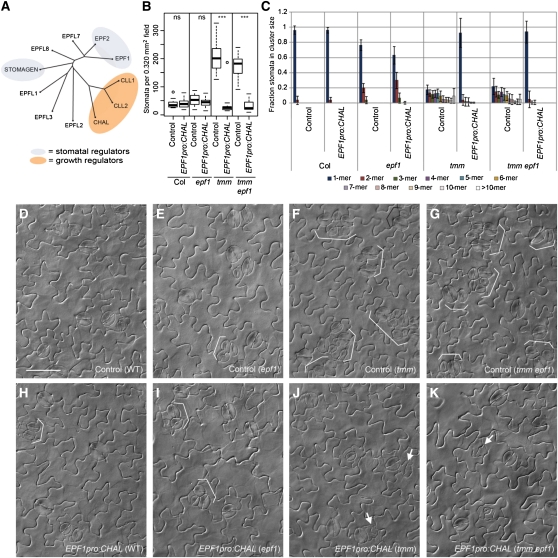
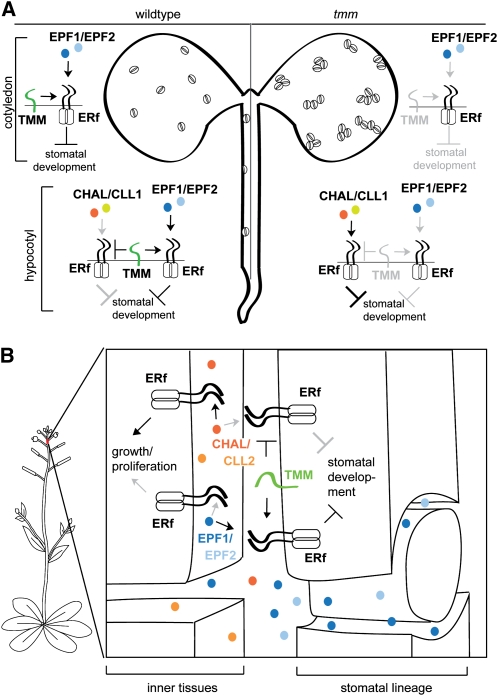
Core signaling pathways function in multiple programs during multicellular development. The mechanisms that compartmentalize pathway function or confer process specificity, however, remain largely unknown. In Arabidopsis thaliana, ERECTA (ER) family receptors have major roles in many growth and cell fate decisions. The ER family acts with receptor TOO MANY MOUTHS (TMM) and several ligands of the EPIDERMAL PATTERNING FACTOR LIKE (EPFL) family, which play distinct yet overlapping roles in patterning of epidermal stomata. Here, our examination of EPFL genes EPFL6/CHALLAH (CHAL), EPFL5/CHALLAH-LIKE1, and EPFL4/CHALLAH-LIKE2 (CLL2) reveals that this family may mediate additional ER-dependent processes. chal cll2 mutants display growth phenotypes characteristic of er mutants, and genetic interactions are consistent with CHAL family molecules acting as ER family ligands. We propose that different classes of EPFL genes regulate different aspects of ER family function and introduce a TMM-based discriminatory mechanism that permits simultaneous, yet compartmentalized and distinct, function of the ER family receptors in growth and epidermal patterning.
Figures
References
-
- Abrash E.B., Bergmann D.C. (2010). Regional specification of stomatal production by the putative ligand CHALLAH. Development 137: 447–455 - PubMed
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Bergmann D.C., Sack F.D. (2007). Stomatal development. Annu. Rev. Plant Biol. 58: 163–181 - PubMed
-
- Bhave N.S., Veley K.M., Nadeau J.A., Lucas J.R., Bhave S.L., Sack F.D. (2009). TOO MANY MOUTHS promotes cell fate progression in stomatal development of Arabidopsis stems. Planta 229: 357–367 - PubMed
-
- Butenko M.A., Vie A.K., Brembu T., Aalen R.B., Bones A.M. (2009). Plant peptides in signalling: Looking for new partners. Trends Plant Sci. 14: 255–263 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
