

Biochemical and genetic requirements for function of the immune response regulator BOTRYTIS-INDUCED KINASE1 in plant growth, ethylene signaling, and PAMP-triggered immunity in Arabidopsis
- PMID: 21862710
- PMCID: PMC3180795
- DOI: 10.1105/tpc.111.087122
Biochemical and genetic requirements for function of the immune response regulator BOTRYTIS-INDUCED KINASE1 in plant growth, ethylene signaling, and PAMP-triggered immunity in Arabidopsis
Abstract
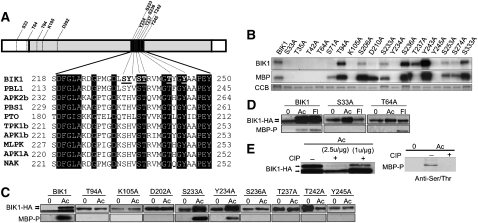
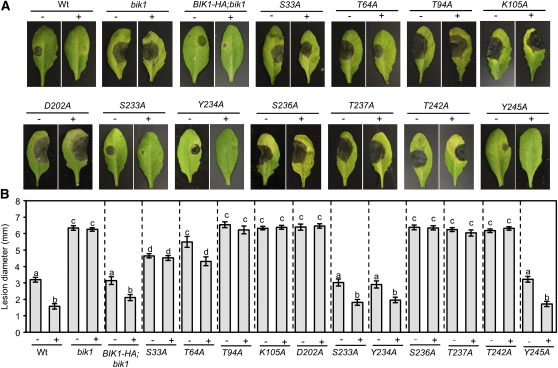
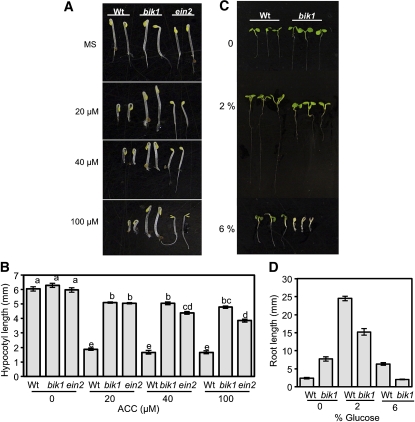
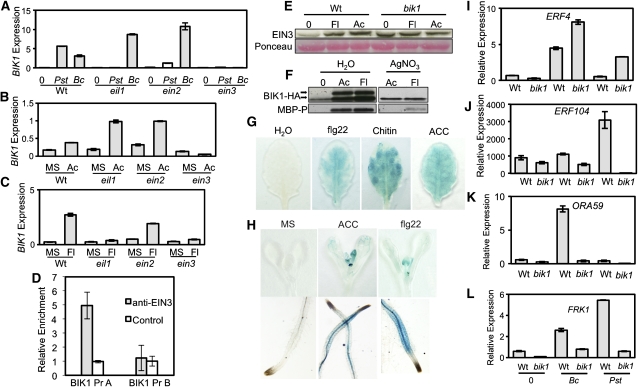
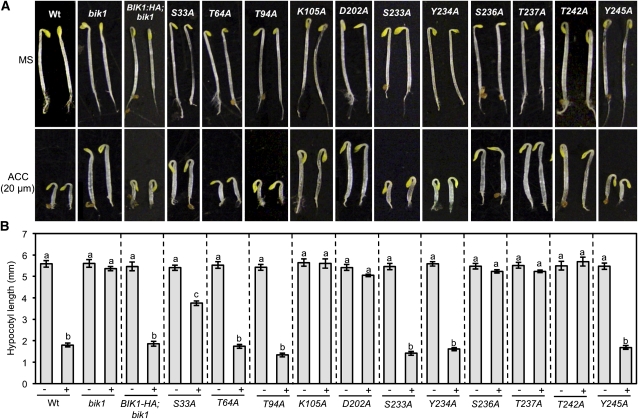
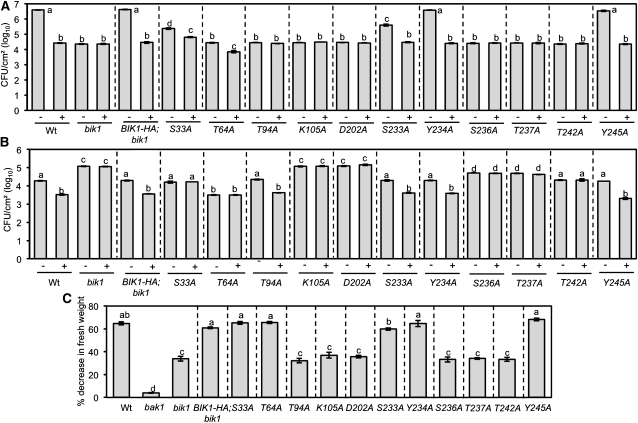
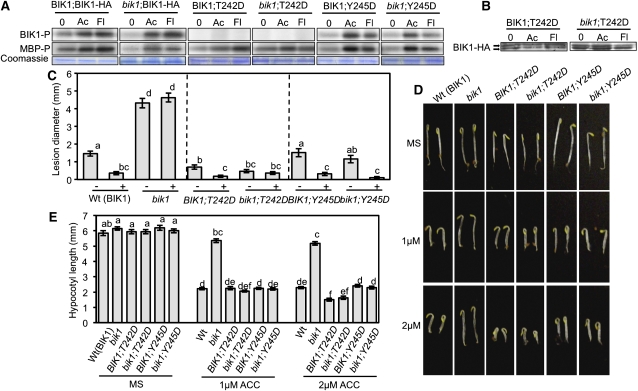
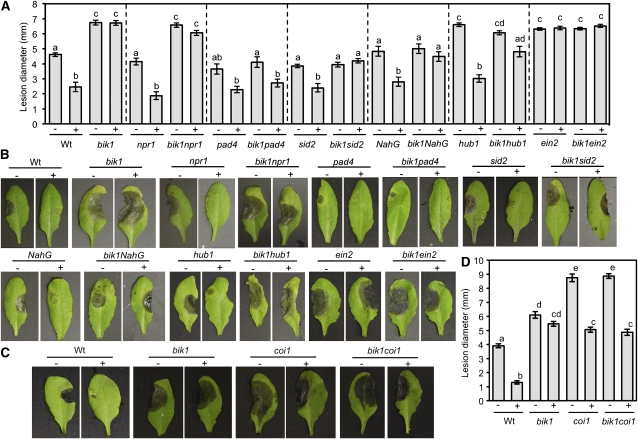
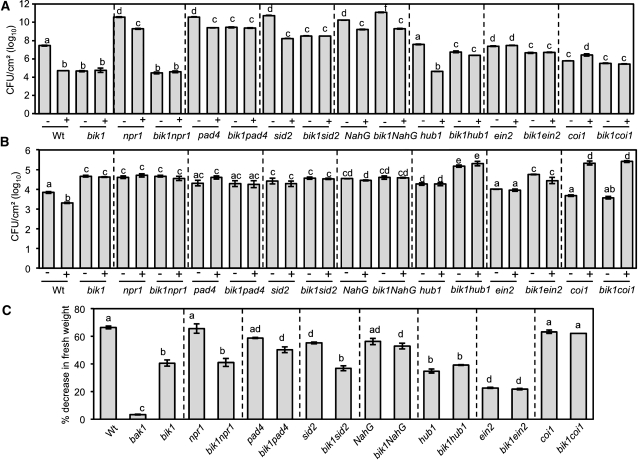
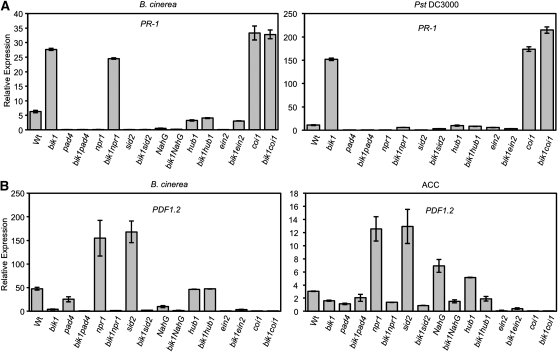
Arabidopsis thaliana BOTRYTIS-INDUCED KINASE1 (BIK1) regulates immune responses to a distinct class of pathogens. Here, mechanisms underlying BIK1 function and its interactions with other immune response regulators were determined. We describe BIK1 function as a component of ethylene (ET) signaling and PAMP-triggered immunity (PTI) to fungal pathogens. BIK1 in vivo kinase activity increases in response to flagellin peptide (flg22) and the ET precursor 1-aminocyclopropane-1-carboxylic acid (ACC) but is blocked by inhibition of ET perception. BIK1 induction by flg22, ACC, and pathogens is strictly dependent on EIN3, and the bik1 mutation results in altered expression of ET-regulated genes. BIK1 site-directed mutants were used to determine residues essential for phosphorylation and biological functions in planta, including PTI, ET signaling, and plant growth. Genetic analysis revealed flg22-induced PTI to Botrytis cinerea requires BIK1, EIN2, and HUB1 but not genes involved in salicylate (SA) functions. BIK1-mediated PTI to Pseudomonas syringae is modulated by SA, ET, and jasmonate signaling. The coi1 mutation suppressed several bik1 phenotypes, suggesting that COI1 may act as a repressor of BIK1 function. Thus, common and distinct mechanisms underlying BIK1 function in mediating responses to distinct pathogens are uncovered. In sum, the critical role of BIK1 in plant immune responses hinges upon phosphorylation, its function in ET signaling, and complex interactions with other immune response regulators.
Figures
Comment in
-
BIK1 function in plant growth and defense signaling.Plant Cell. 2011 Aug;23(8):2806. doi: 10.1105/tpc.111.230811. Epub 2011 Aug 23. Plant Cell. 2011. PMID: 21862709 Free PMC article. No abstract available.
Similar articles
-
The Multifaceted Ubiquitination of BIK1 During Plant Immunity in Arabidopsis thaliana.Int J Mol Sci. 2024 Nov 13;25(22):12187. doi: 10.3390/ijms252212187. Int J Mol Sci. 2024. PMID: 39596247 Free PMC article. Review.
-
ERF5 and ERF6 play redundant roles as positive regulators of JA/Et-mediated defense against Botrytis cinerea in Arabidopsis.PLoS One. 2012;7(4):e35995. doi: 10.1371/journal.pone.0035995. Epub 2012 Apr 26. PLoS One. 2012. PMID: 22563431 Free PMC article.
-
The receptor-like cytoplasmic kinase PCRK1 contributes to pattern-triggered immunity against Pseudomonas syringae in Arabidopsis thaliana.New Phytol. 2015 Jul;207(1):78-90. doi: 10.1111/nph.13345. Epub 2015 Feb 25. New Phytol. 2015. PMID: 25711411
-
Bacillus cereus AR156 Activates Defense Responses to Pseudomonas syringae pv. tomato in Arabidopsis thaliana Similarly to flg22.Mol Plant Microbe Interact. 2018 Mar;31(3):311-322. doi: 10.1094/MPMI-10-17-0240-R. Epub 2018 Jan 8. Mol Plant Microbe Interact. 2018. PMID: 29090631
-
Activation and turnover of the plant immune signaling kinase BIK1: a fine balance.Essays Biochem. 2022 Aug 5;66(2):207-218. doi: 10.1042/EBC20210071. Essays Biochem. 2022. PMID: 35575190 Review.
Cited by
-
Combined roles of ethylene and endogenous peptides in regulating plant immunity and growth.Proc Natl Acad Sci U S A. 2013 Apr 9;110(15):5748-9. doi: 10.1073/pnas.1302659110. Epub 2013 Mar 25. Proc Natl Acad Sci U S A. 2013. PMID: 23530253 Free PMC article. No abstract available.
-
Differential response of tomato genotypes to Xanthomonas-specific pathogen-associated molecular patterns and correlation with bacterial spot (Xanthomonas perforans) resistance.Hortic Res. 2016 Aug 10;3:16035. doi: 10.1038/hortres.2016.35. eCollection 2016. Hortic Res. 2016. PMID: 27555919 Free PMC article.
-
The Receptor-like Cytoplasmic Kinase BIK1 Localizes to the Nucleus and Regulates Defense Hormone Expression during Plant Innate Immunity.Cell Host Microbe. 2018 Apr 11;23(4):485-497.e5. doi: 10.1016/j.chom.2018.03.010. Cell Host Microbe. 2018. PMID: 29649442 Free PMC article.
-
BOTRYTIS-INDUCED KINASE1 Modulates Arabidopsis Resistance to Green Peach Aphids via PHYTOALEXIN DEFICIENT4.Plant Physiol. 2014 Aug;165(4):1657-1670. doi: 10.1104/pp.114.242206. Epub 2014 Jun 24. Plant Physiol. 2014. PMID: 24963070 Free PMC article.
-
The Multifaceted Ubiquitination of BIK1 During Plant Immunity in Arabidopsis thaliana.Int J Mol Sci. 2024 Nov 13;25(22):12187. doi: 10.3390/ijms252212187. Int J Mol Sci. 2024. PMID: 39596247 Free PMC article. Review.
References
-
- Becraft P.W. (2002). Receptor kinase signaling in plant development. Annu. Rev. Cell Dev. Biol. 18: 163–192 - PubMed
-
- Boller T., Felix G. (2009). A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60: 379–406 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
