

Active sampling and decision making in Drosophila chemotaxis
- PMID: 21863008
- PMCID: PMC3265367
- DOI: 10.1038/ncomms1455
Active sampling and decision making in Drosophila chemotaxis
Abstract
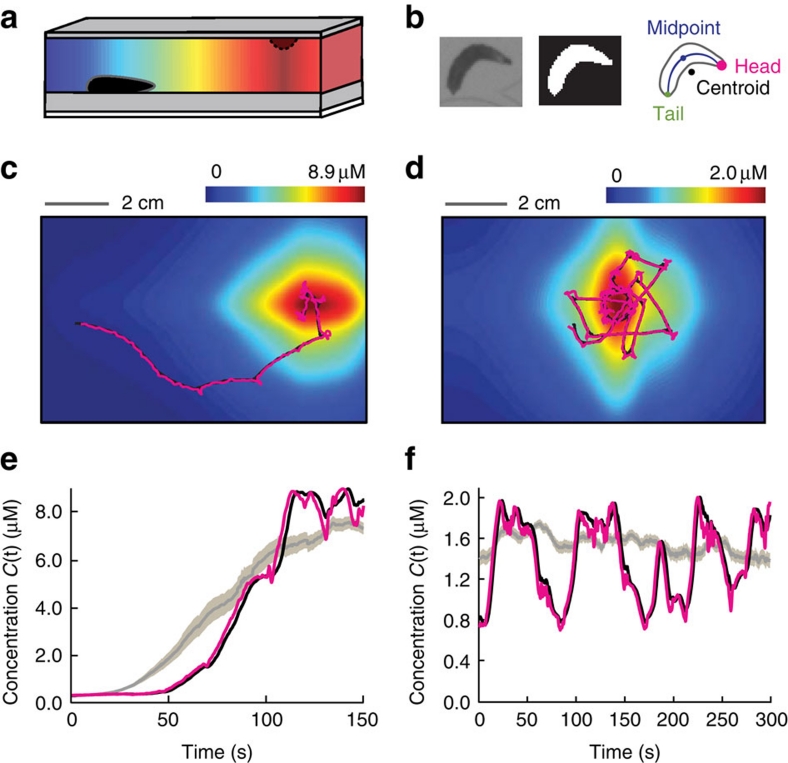
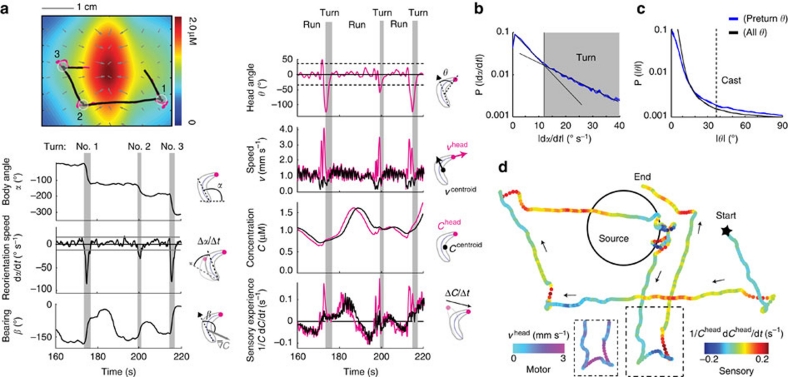
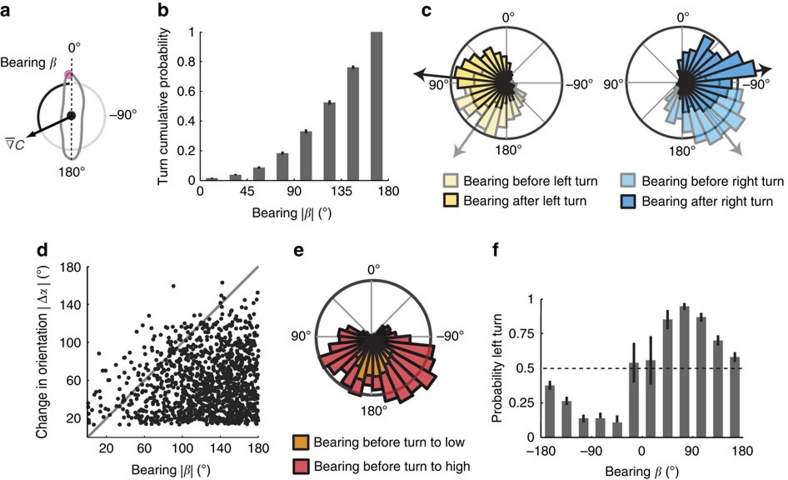
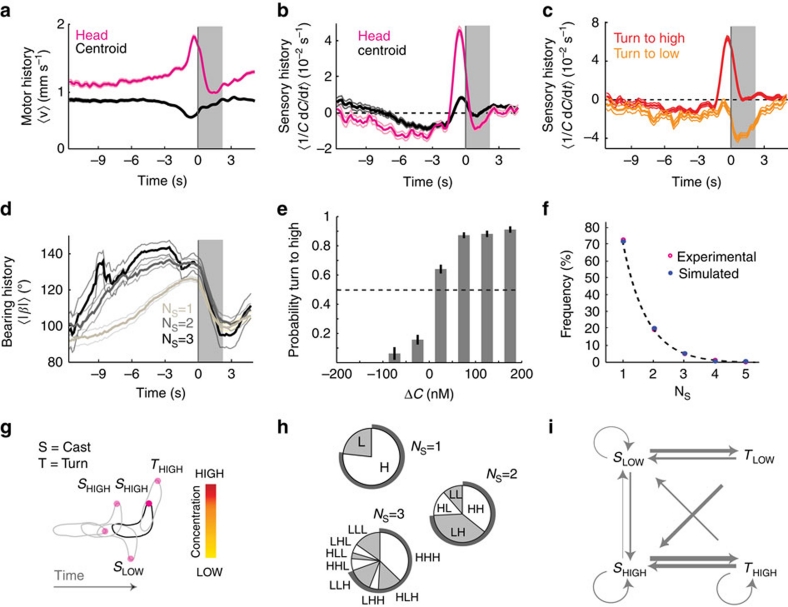
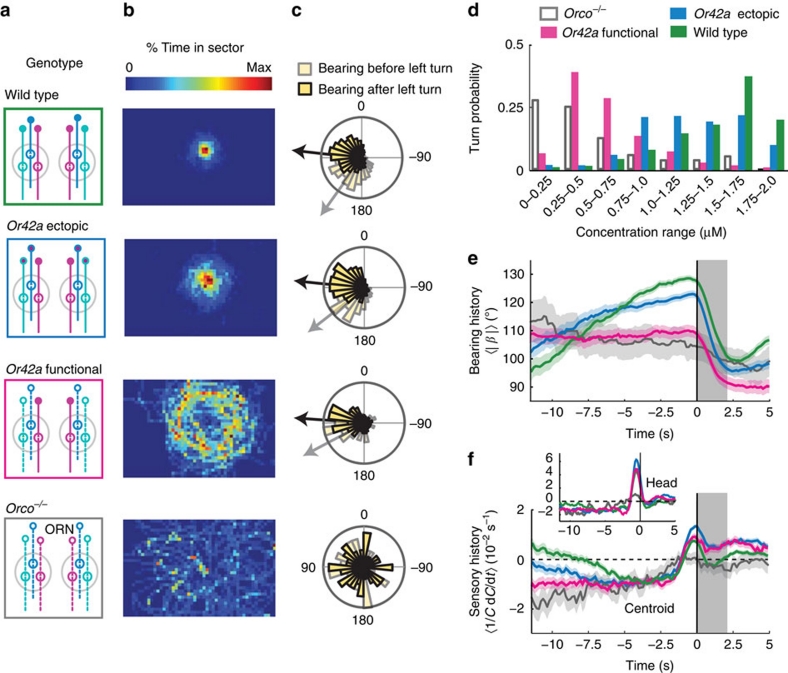
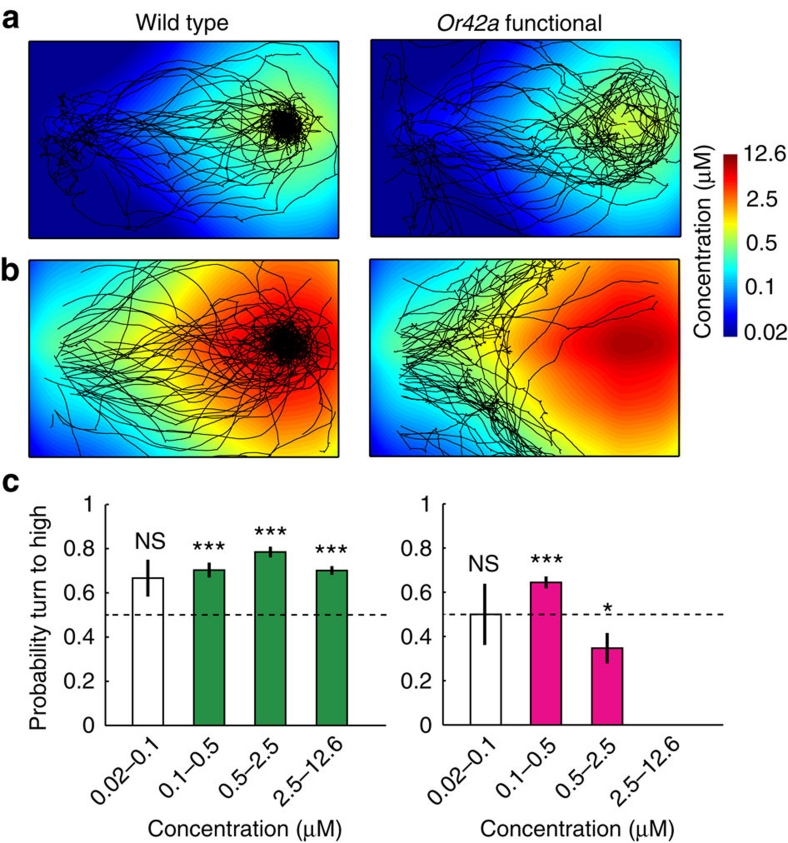
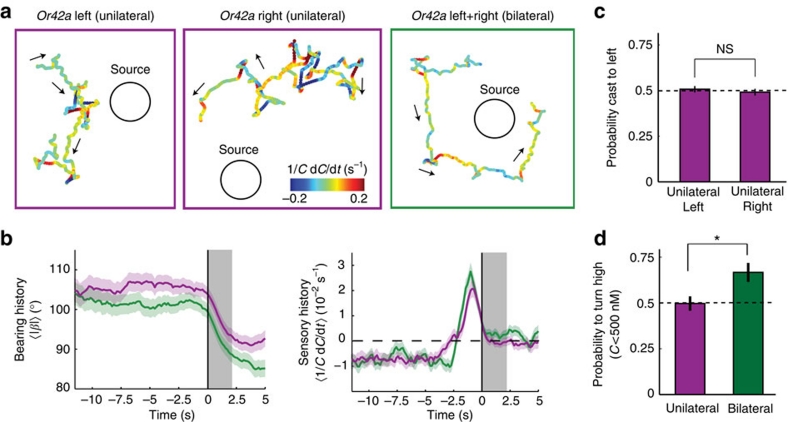
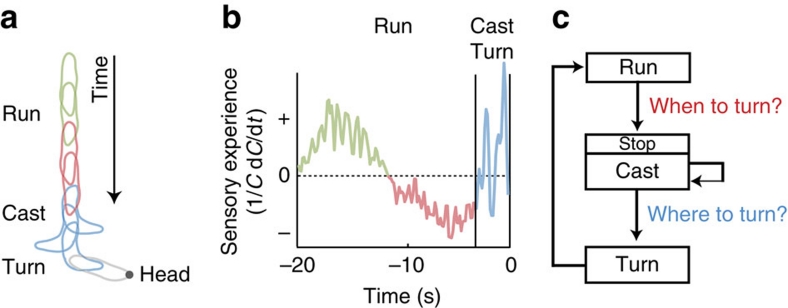
The ability to respond to chemical stimuli is fundamental to the survival of motile organisms, but the strategies underlying odour tracking remain poorly understood. Here we show that chemotaxis in Drosophila melanogaster larvae is an active sampling process analogous to sniffing in vertebrates. Combining computer-vision algorithms with reconstructed olfactory environments, we establish that larvae orient in odour gradients through a sequential organization of stereotypical behaviours, including runs, stops, lateral head casts and directed turns. Negative gradients, integrated during runs, control the timing of turns. Positive gradients detected through high-amplitude head casts determine the direction of individual turns. By genetically manipulating the peripheral olfactory circuit, we examine how orientation adapts to losses and gains of function in olfactory input. Our findings suggest that larval chemotaxis represents an intermediate navigation strategy between the biased random walks of Escherichia Coli and the stereo-olfaction observed in rats and humans.
© 2011 Macmillan Publishers Limited. All rights reserved.
Figures
References
-
- Berg H. C. Motile behavior of bacteria. Phys. Today 53, 24–29 (2000).
-
- Thesen A., Steen J. B. & Doving K. B. Behaviour of dogs during olfactory tracking. J. Exp. Biol. 180, 247–251 (1993). - PubMed
-
- Rajan R., Clement J. P. & Bhalla U. S. Rats smell in stereo. Science 311, 666–670 (2006). - PubMed
-
- Porter J. et al. Mechanisms of scent-tracking in humans. Nat. Neurosci. 10, 27–29 (2007). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
