

Vav protein guanine nucleotide exchange factor regulates CD36 protein-mediated macrophage foam cell formation via calcium and dynamin-dependent processes
- PMID: 21865158
- PMCID: PMC3195569
- DOI: 10.1074/jbc.M111.265082
Vav protein guanine nucleotide exchange factor regulates CD36 protein-mediated macrophage foam cell formation via calcium and dynamin-dependent processes
Abstract
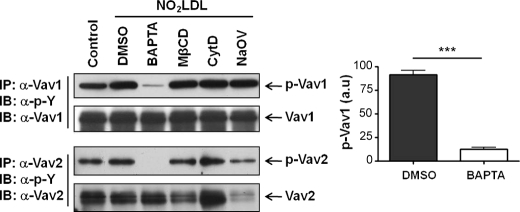
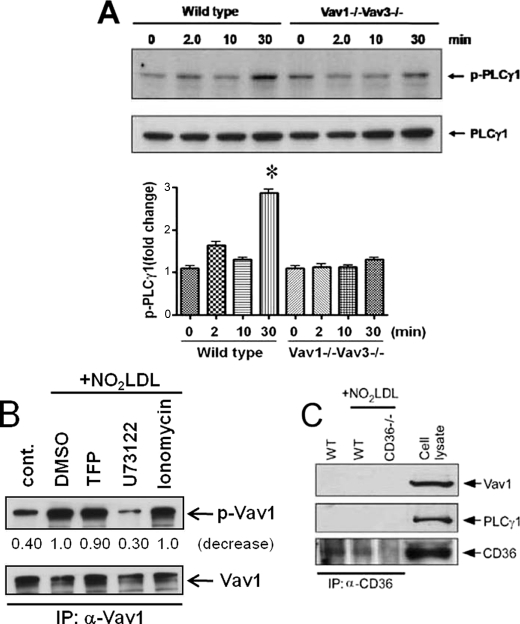
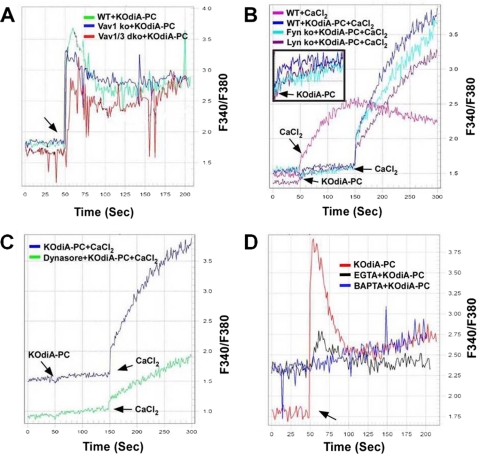
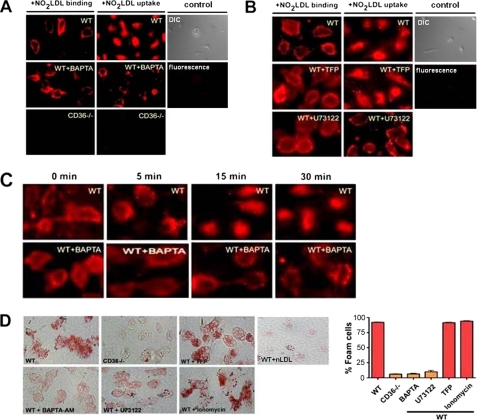
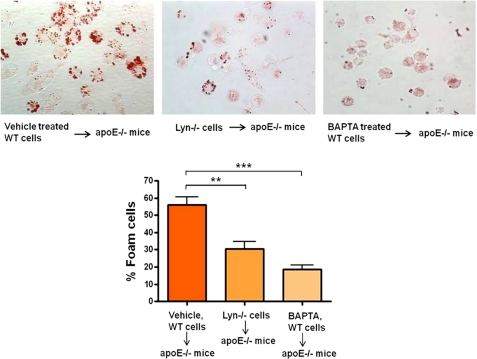
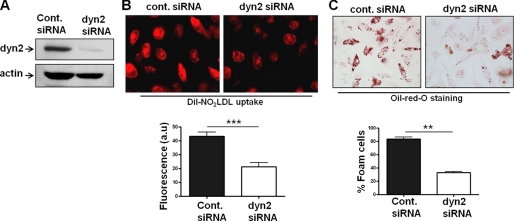
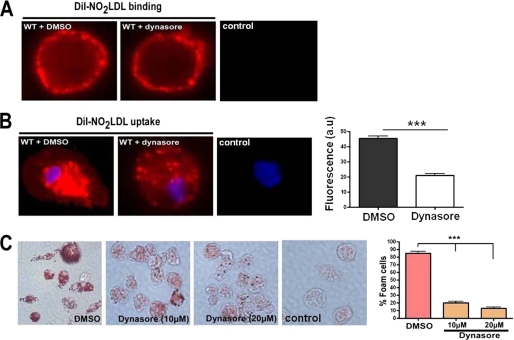
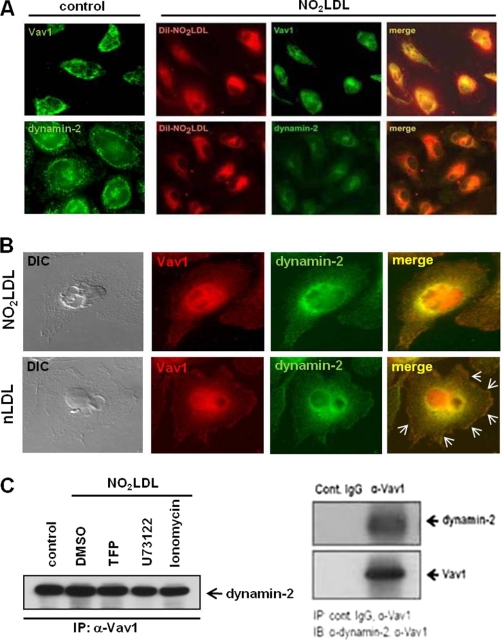
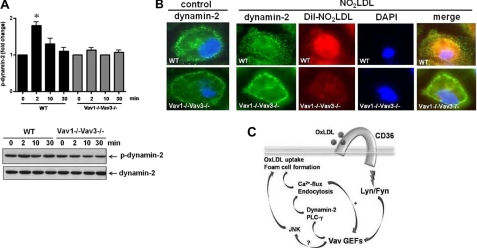
Atherosclerosis, a chronic inflammatory disease, results in part from the accumulation of modified lipoproteins in the arterial wall and formation of lipid-laden macrophages, known as "foam cells." Recently, we reported that CD36, a scavenger receptor, contributes to activation of Vav-family guanine nucleotide exchange factors by oxidatively modified LDL in macrophages. We also discovered that CD36-dependent uptake of oxidized LDL (oxLDL) in vitro and foam cell formation in vitro and in vivo was significantly reduced in macrophages deficient of Vav proteins. The goal of the present study was to identify the mechanisms by which Vav proteins regulate CD36-dependent foam cell formation. We now show that a Vav-dynamin signaling axis plays a critical role in generating calcium signals in mouse macrophages exposed to CD36-specific oxidized phospholipid ligands. Chelation of intracellular Ca(2+) or inhibition of phospholipase C-γ (PLC-γ) inhibited Vav activation (85 and 70%, respectively, compared with vehicle control) and reduced foam cell formation (approximately 75%). Knockdown of expression by siRNA or inhibition of GTPase activity of dynamin 2, a Vav-interacting protein involved in endocytic vesicle fission, significantly blocked oxLDL uptake and inhibited foam cell formation. Immunofluorescence microscopy studies showed that Vav1 and dynamin 2 colocalized with internalized oxLDL in macrophages and that activation and mobilization of dynamin 2 by oxLDL was impaired in vav null cells. These studies identified previously unknown components of the CD36 signaling pathway, demonstrating that Vav proteins regulate oxLDL uptake and foam cell formation via calcium- and dynamin 2-dependent processes and thus represent novel therapeutic targets for atherosclerosis.
Figures
Similar articles
-
Nicotine potentiates proatherogenic effects of oxLDL by stimulating and upregulating macrophage CD36 signaling.Am J Physiol Heart Circ Physiol. 2013 Aug 15;305(4):H563-74. doi: 10.1152/ajpheart.00042.2013. Epub 2013 Jun 7. Am J Physiol Heart Circ Physiol. 2013. PMID: 23748423 Free PMC article.
-
Vav family Rho guanine nucleotide exchange factors regulate CD36-mediated macrophage foam cell formation.J Biol Chem. 2011 Mar 4;286(9):7010-7. doi: 10.1074/jbc.M110.192450. Epub 2011 Jan 5. J Biol Chem. 2011. PMID: 21209086 Free PMC article.
-
Vav Guanine nucleotide exchange factors regulate atherosclerotic lesion development in mice.Arterioscler Thromb Vasc Biol. 2013 Sep;33(9):2053-7. doi: 10.1161/ATVBAHA.113.301414. Epub 2013 Jul 3. Arterioscler Thromb Vasc Biol. 2013. PMID: 23825362 Free PMC article.
-
Mechanisms of cell signaling by the scavenger receptor CD36: implications in atherosclerosis and thrombosis.Trans Am Clin Climatol Assoc. 2010;121:206-20. Trans Am Clin Climatol Assoc. 2010. PMID: 20697562 Free PMC article. Review.
-
Foam cells in atherosclerosis.Clin Chim Acta. 2013 Sep 23;424:245-52. doi: 10.1016/j.cca.2013.06.006. Epub 2013 Jun 16. Clin Chim Acta. 2013. PMID: 23782937 Review.
Cited by
-
Lipoprotein(a) the Insurgent: A New Insight into the Structure, Function, Metabolism, Pathogenicity, and Medications Affecting Lipoprotein(a) Molecule.J Lipids. 2020 Feb 1;2020:3491764. doi: 10.1155/2020/3491764. eCollection 2020. J Lipids. 2020. PMID: 32099678 Free PMC article. Review.
-
Oxidized LDL-bound CD36 recruits an Na⁺/K⁺-ATPase-Lyn complex in macrophages that promotes atherosclerosis.Sci Signal. 2015 Sep 8;8(393):ra91. doi: 10.1126/scisignal.aaa9623. Sci Signal. 2015. PMID: 26350901 Free PMC article.
-
The computational analyses, molecular dynamics of fatty-acid transport mechanism to the CD36 receptor.Sci Rep. 2021 Dec 1;11(1):23207. doi: 10.1038/s41598-021-01373-5. Sci Rep. 2021. PMID: 34853341 Free PMC article.
-
Nicotine potentiates proatherogenic effects of oxLDL by stimulating and upregulating macrophage CD36 signaling.Am J Physiol Heart Circ Physiol. 2013 Aug 15;305(4):H563-74. doi: 10.1152/ajpheart.00042.2013. Epub 2013 Jun 7. Am J Physiol Heart Circ Physiol. 2013. PMID: 23748423 Free PMC article.
-
Cannabis sativa extracts inhibit LDL oxidation and the formation of foam cells in vitro, acting as potential multi-step inhibitors of atherosclerosis development.PLoS One. 2024 Dec 20;19(12):e0310777. doi: 10.1371/journal.pone.0310777. eCollection 2024. PLoS One. 2024. PMID: 39705234 Free PMC article.
References
-
- Glass C. K., Witztum J. L. (2001) Cell 104, 503–516 - PubMed
-
- Podrez E. A., Poliakov E., Shen Z., Zhang R., Deng Y., Sun M., Finton P. J., Shan L., Febbraio M., Hajjar D. P., Silverstein R. L., Hoff H. F., Salomon R. G., Hazen S. L. (2002) J. Biol. Chem. 277, 38517–38523 - PubMed
-
- Zhao Z., de Beer M. C., Cai L., Asmis R., de Beer F. C., de Villiers W. J., van der, Westhuyzen D. R. (2005) Arterioscler. Thromb. Vasc. Biol. 25, 168–173 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
