

Tilted cellulose arrangement as a novel mechanism for hygroscopic coiling in the stork's bill awn
- PMID: 21865252
- PMCID: PMC3284132
- DOI: 10.1098/rsif.2011.0395
Tilted cellulose arrangement as a novel mechanism for hygroscopic coiling in the stork's bill awn
Abstract
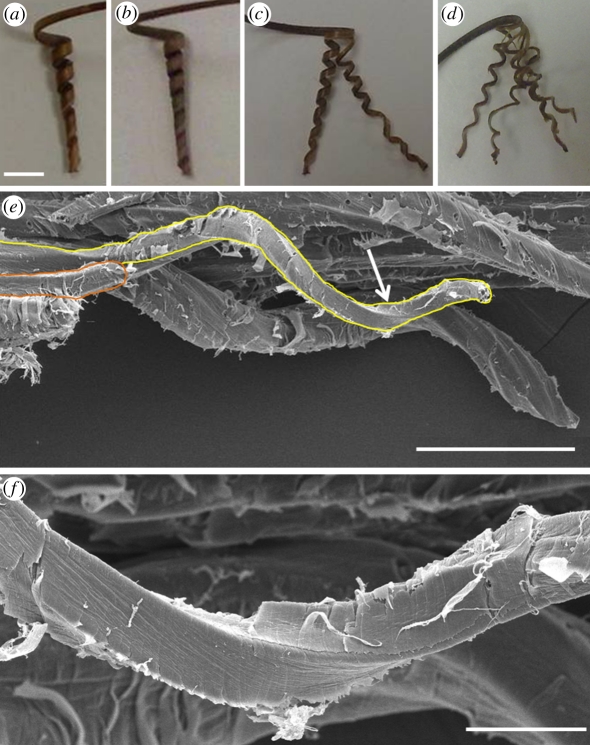
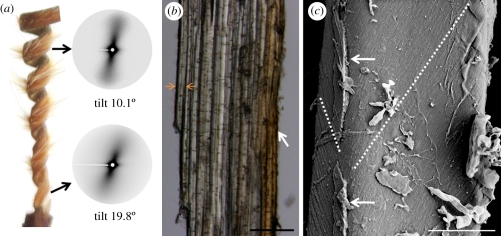
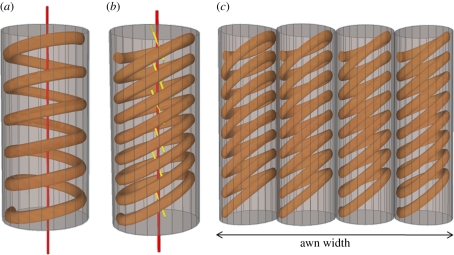
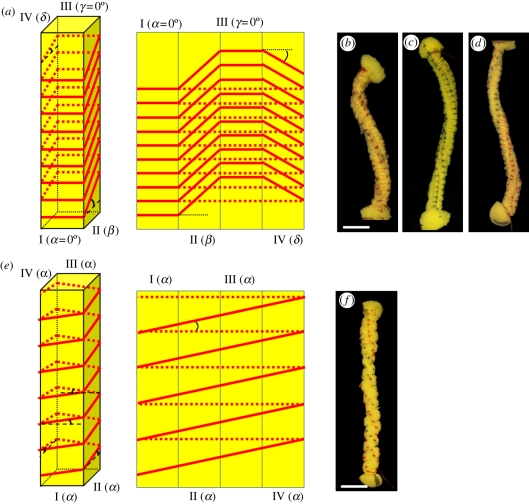
The sessile nature of plants demands the development of seed-dispersal mechanisms to establish new growing loci. Dispersal strategies of many species involve drying of the dispersal unit, which induces directed contraction and movement based on changing environmental humidity. The majority of researched hygroscopic dispersal mechanisms are based on a bilayered structure. Here, we investigate the motility of the stork's bill (Erodium) seeds that relies on the tightening and loosening of a helical awn to propel itself across the surface into a safe germination place. We show that this movement is based on a specialized single layer consisting of a mechanically uniform tissue. A cell wall structure with cellulose microfibrils arranged in an unusually tilted helix causes each cell to spiral. These cells generate a macroscopic coil by spiralling collectively. A simple model made from a thread embedded in an isotropic foam matrix shows that this cellulose arrangement is indeed sufficient to induce the spiralling of the cells.
Figures


References
-
- Fowler N. L. 1988. What is a safe site? Neighbor, litter, germination date, and patch effects. Ecology 69, 947–961 10.2307/1941250 (doi:10.2307/1941250) - DOI
-
- Van der Pijl L. 1982. Principles of dispersal in higher plants, 3rd edn. New York, NY: Springer
-
- Fahn A., Werker E. 1972. Anatomical mechanisms of seed dispersal. In Seed biology (ed. Kozlowski T. T.), pp. 151–221 New York, NY: Academic Press
-
- Harper J. L., Lovell P. H., Moore K. G. 1970. The shapes and sizes of seeds. Annu. Rev. Ecol. Syst. 1, 327–356 10.1146/annurev.es.01.110170.001551 (doi:10.1146/annurev.es.01.110170.001551) - DOI
-
- Elbaum R., Zaltzman L., Burgert I., Fratzl P. 2007. The role of wheat awns in the seed dispersal unit. Science 316, 884–886 10.1126/science.1140097 (doi:10.1126/science.1140097) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources