

Dynamic antibody specificities and virion concentrations in circulating immune complexes in acute to chronic HIV-1 infection
- PMID: 21865397
- PMCID: PMC3194959
- DOI: 10.1128/JVI.05601-11
Dynamic antibody specificities and virion concentrations in circulating immune complexes in acute to chronic HIV-1 infection
Abstract
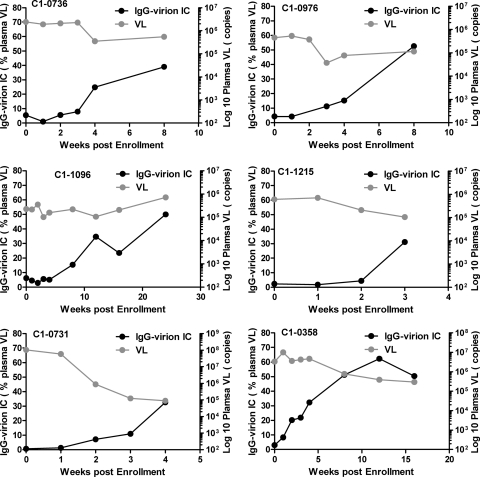
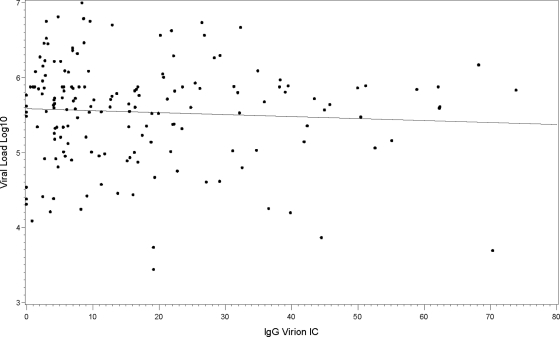
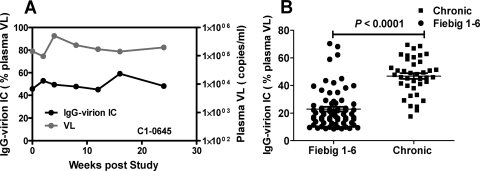
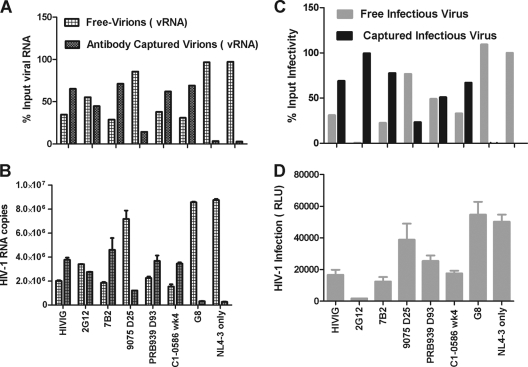
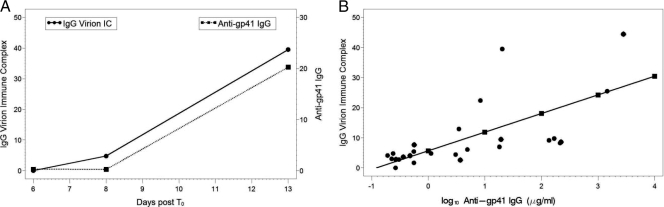
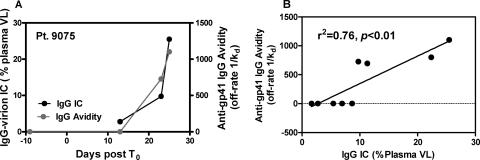
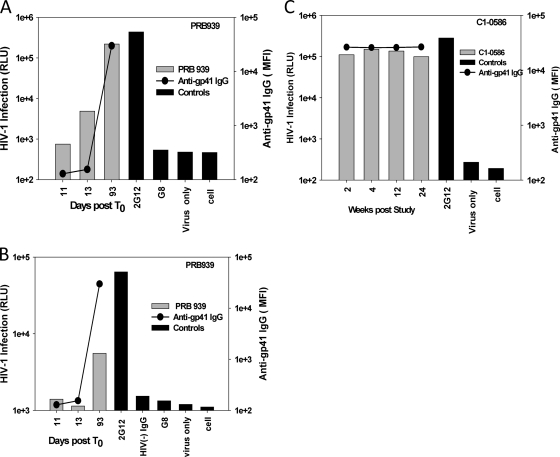
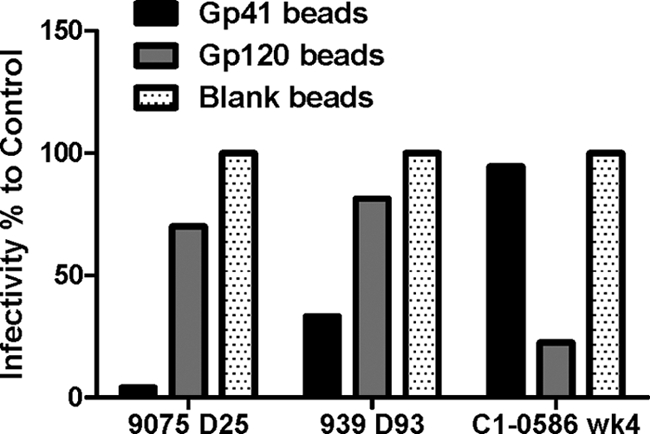
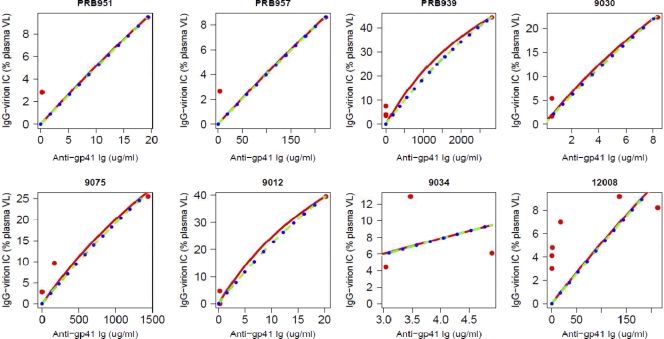
Understanding the interactions between human immunodeficiency virus type 1 (HIV-1) virions and antibodies (Ab) produced during acute HIV-1 infection (AHI) is critical for defining antibody antiviral capabilities. Antibodies that bind virions may prevent transmission by neutralization of virus or mechanically prevent HIV-1 migration through mucosal layers. In this study, we quantified circulating HIV-1 virion-immune complexes (ICs), present in approximately 90% of AHI subjects, and compared the levels and antibody specificity to those in chronic infection. Circulating HIV-1 virions coated with IgG (immune complexes) were in significantly lower levels relative to the viral load in acute infection than in chronic HIV-1 infection. The specificities of the antibodies in the immune complexes differed between acute and chronic infection (anti-gp41 Ab in acute infection and anti-gp120 in chronic infection), potentially suggesting different roles in immunopathogenesis for complexes arising at different stages of infection. We also determined the ability of circulating IgG from AHI to bind infectious versus noninfectious virions. Similar to a nonneutralizing anti-gp41 monoclonal antibody (MAb), purified plasma IgG from acute HIV-1 subjects bound both infectious and noninfectious virions. This was in contrast to the neutralizing antibody 2G12 MAb that bound predominantly infectious virions. Moreover, the initial antibody response captured acute HIV-1 virions without selection for different HIV-1 envelope sequences. In total, this study demonstrates that the composition of immune complexes are dynamic over the course of HIV-1 infection and are comprised initially of antibodies that nonselectively opsonize both infectious and noninfectious virions, likely contributing to the lack of efficacy of the antibody response during acute infection.
Figures
References
-
- Bajtay Z., Speth C., Erdei A., Dierich M. P. 2004. Cutting edge: productive HIV-1 infection of dendritic cells via complement receptor type 3 (CR3, CD11b/CD18). J. Immunol. 173:4775–4778 - PubMed
-
- Bouhlal H., et al. 2002. Opsonization of HIV-1 by semen complement enhances infection of human epithelial cells. J. Immunol. 169:3301–3306 - PubMed
-
- Bourinbaiar A. S. 1994. The ratio of defective HIV-1 particles to replication-competent infectious virions. Acta Virol. 38:59–61 - PubMed
-
- Burrer R., Haessig-Einius S., Aubertin A. M., Moog C. 2005. Neutralizing as well as non-neutralizing polyclonal immunoglobulin (Ig)G from infected patients capture HIV-1 via antibodies directed against the principal immunodominant domain of gp41. Virology 333:102–113 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
