

Single synapse information coding in intraburst spike patterns of central pattern generator motor neurons
- PMID: 21865472
- PMCID: PMC6623235
- DOI: 10.1523/JNEUROSCI.1568-11.2011
Single synapse information coding in intraburst spike patterns of central pattern generator motor neurons
Abstract
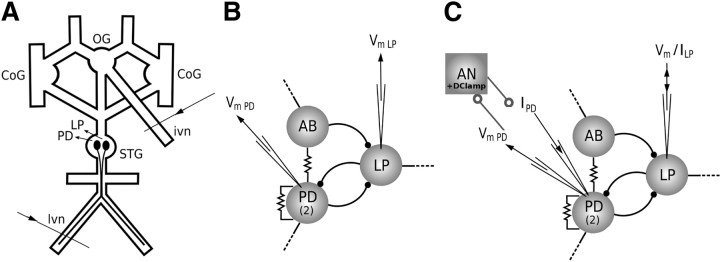
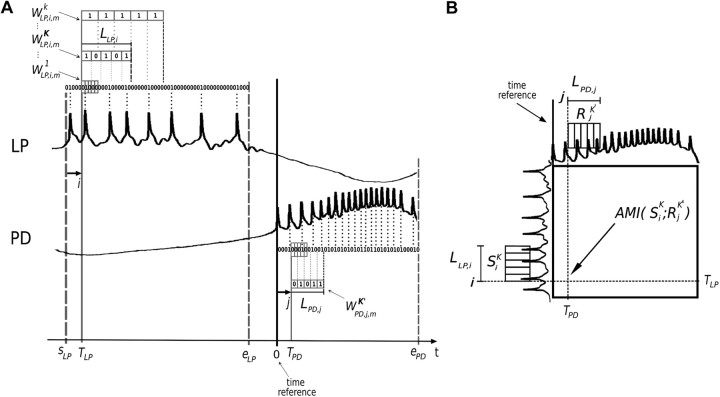
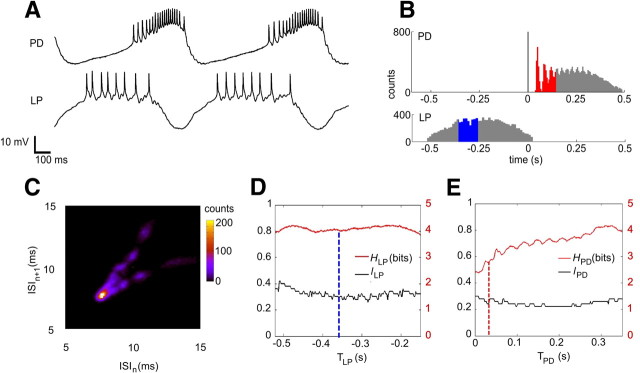
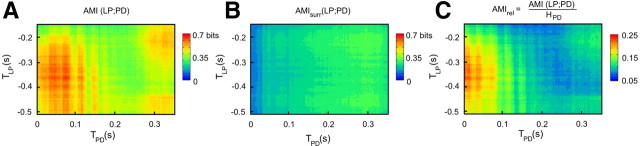
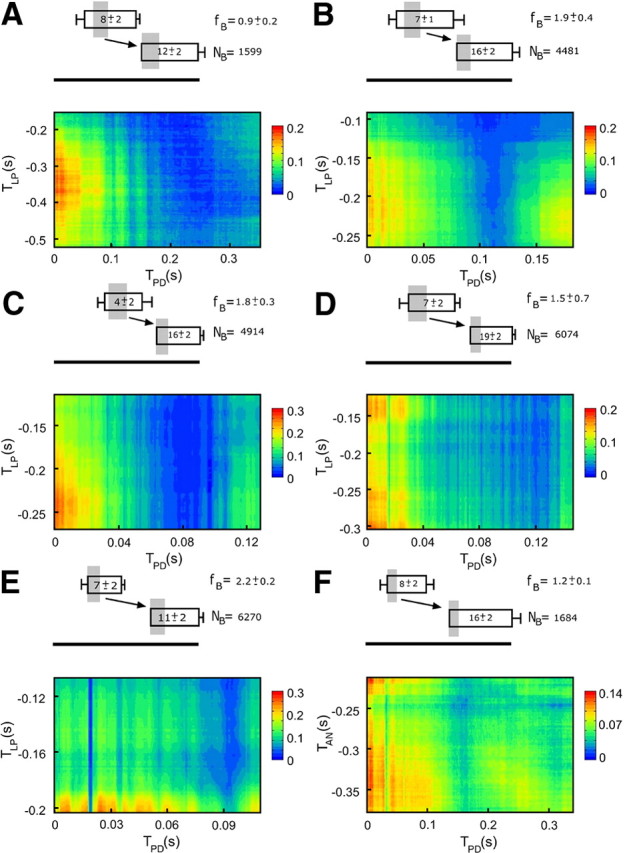
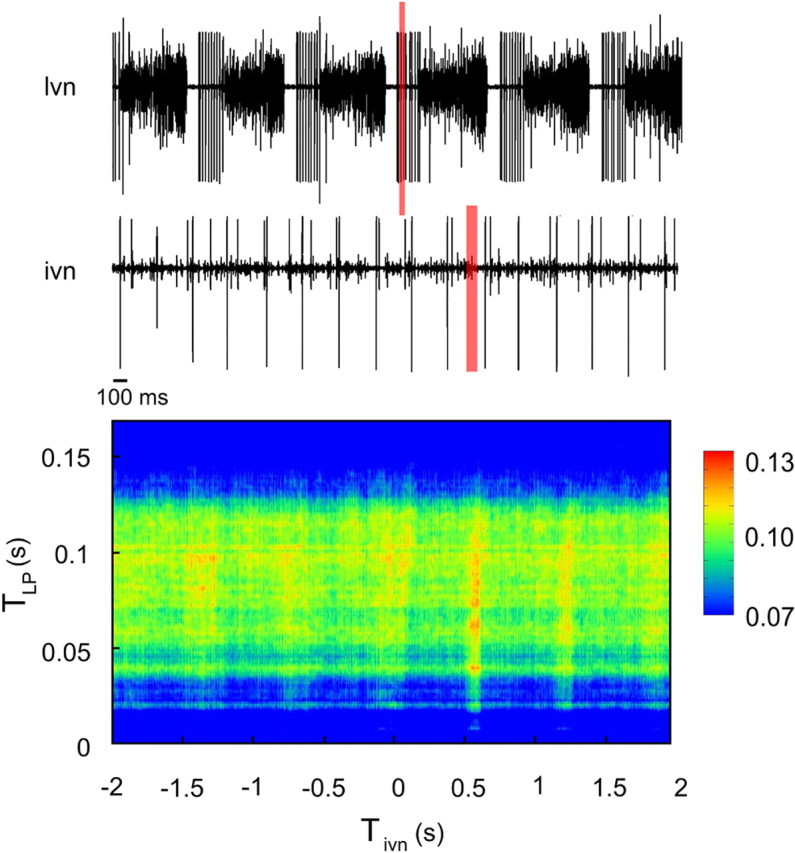
Burst firing is ubiquitous in nervous systems and has been intensively studied in central pattern generators (CPGs). Previous works have described subtle intraburst spike patterns (IBSPs) that, despite being traditionally neglected for their lack of relation to CPG motor function, were shown to be cell-type specific and sensitive to CPG connectivity. Here we address this matter by investigating how a bursting motor neuron expresses information about other neurons in the network. We performed experiments on the crustacean stomatogastric pyloric CPG, both in control conditions and interacting in real-time with computer model neurons. The sensitivity of postsynaptic to presynaptic IBSPs was inferred by computing their average mutual information along each neuron burst. We found that details of input patterns are nonlinearly and inhomogeneously coded through a single synapse into the fine IBSPs structure of the postsynaptic neuron following burst. In this way, motor neurons are able to use different time scales to convey two types of information simultaneously: muscle contraction (related to bursting rhythm) and the behavior of other CPG neurons (at a much shorter timescale by using IBSPs as information carriers). Moreover, the analysis revealed that the coding mechanism described takes part in a previously unsuspected information pathway from a CPG motor neuron to a nerve that projects to sensory brain areas, thus providing evidence of the general physiological role of information coding through IBSPs in the regulation of neuronal firing patterns in remote circuits by the CNS.
Figures
References
-
- Ayers J, Selverston AI. Synaptic perturbation and entrainment of gastric mill rhythm of the spiny lobster. J Neurophysiol. 1984;51:113–125. - PubMed
-
- Bair W, Koch C. Temporal precision of spike trains in extrastriate cortex of the behaving macaque monkey. Neural Comput. 1996;8:1185–1202. - PubMed
-
- Böhm H, Dybek E, Heizel HG. Anatomy and in vivo activity of neurons connecting the crustacean stomatogastric nervous system to the brain. J Comp Physiol A. 2001;187:392–403. - PubMed
-
- Borst A, Theunissen FE. Information theory and neural coding. Nat Neurosci. 1999;2:947–957. - PubMed
-
- Brocard F, Tazerart S, Vinay L. Do pacemakers drive the central pattern generator for locomotion in mammals? Neuroscientist. 2010;16:139–155. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources