

Parkin is a lipid-responsive regulator of fat uptake in mice and mutant human cells
- PMID: 21865652
- PMCID: PMC3171101
- DOI: 10.1172/JCI44736
Parkin is a lipid-responsive regulator of fat uptake in mice and mutant human cells
Abstract
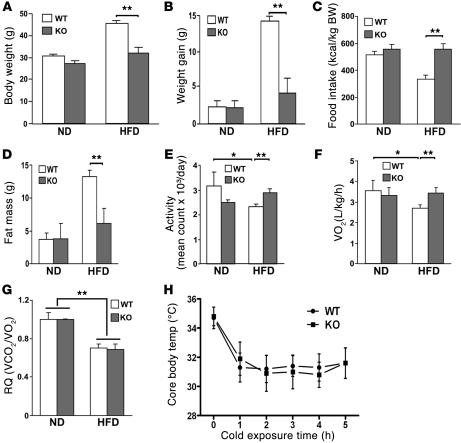
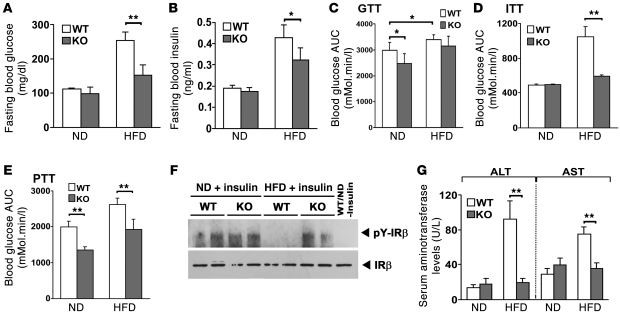
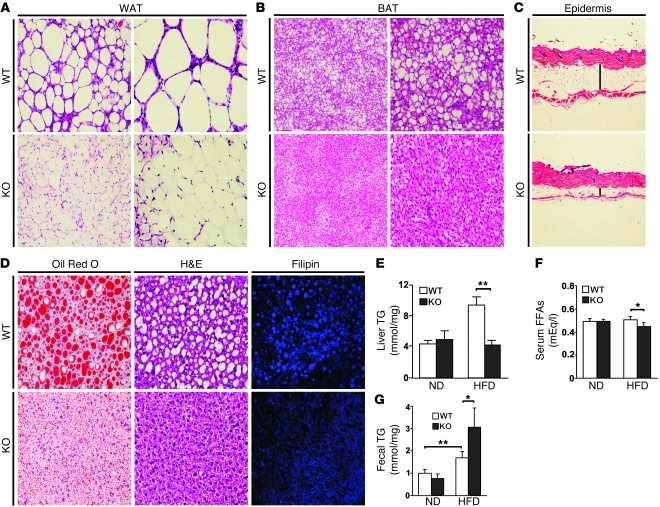
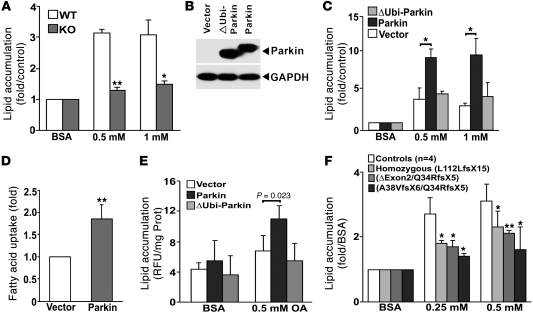
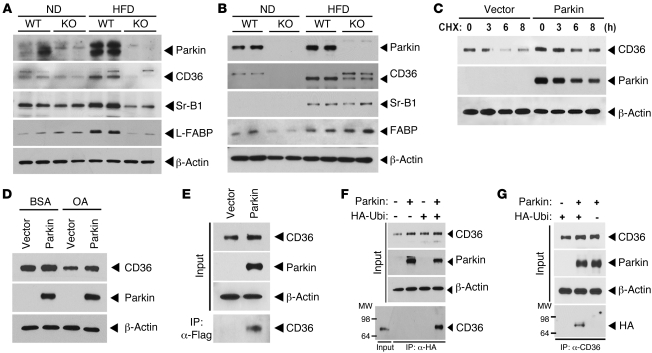
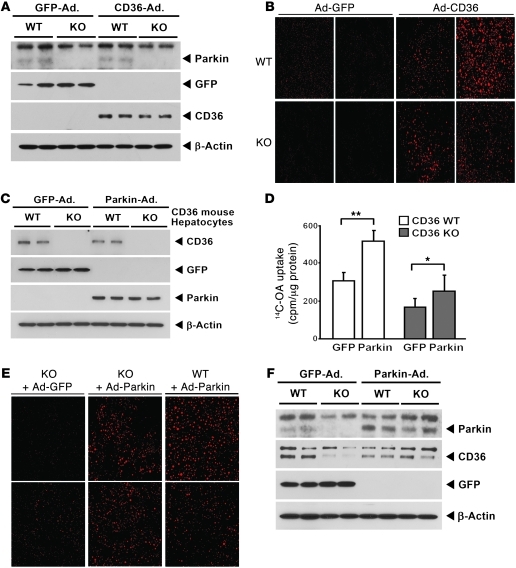
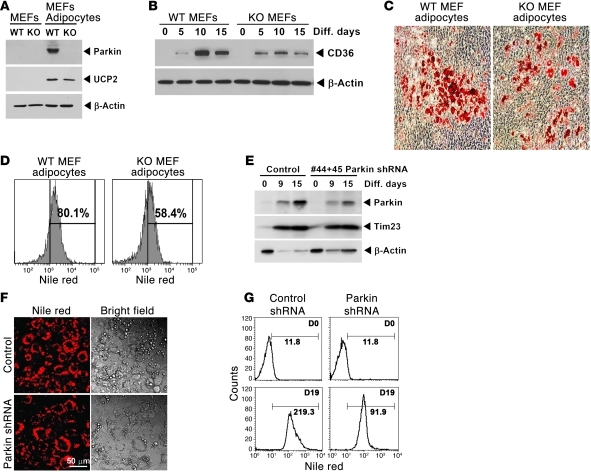
It has long been hypothesized that abnormalities in lipid biology contribute to degenerative brain diseases. Consistent with this, emerging epidemiologic evidence links lipid alterations with Parkinson disease (PD), and disruption of lipid metabolism has been found to predispose to α-synuclein toxicity. We therefore investigated whether Parkin, an E3 ubiquitin ligase found to be defective in patients with early onset PD, regulates systemic lipid metabolism. We perturbed lipid levels by exposing Parkin+/+ and Parkin-/- mice to a high-fat and -cholesterol diet (HFD). Parkin-/- mice resisted weight gain, steatohepatitis, and insulin resistance. In wild-type mice, the HFD markedly increased hepatic Parkin levels in parallel with lipid transport proteins, including CD36, Sr-B1, and FABP. These lipid transport proteins were not induced in Parkin-/- mice. The role of Parkin in fat uptake was confirmed by increased oleate accumulation in hepatocytes overexpressing Parkin and decreased uptake in Parkin-/- mouse embryonic fibroblasts and patient cells harboring complex heterozygous mutations in the Parkin-encoding gene PARK2. Parkin conferred this effect, in part, via ubiquitin-mediated stabilization of the lipid transporter CD36. Reconstitution of Parkin restored hepatic fat uptake and CD36 levels in Parkin-/- mice, and Parkin augmented fat accumulation during adipocyte differentiation. These results demonstrate that Parkin is regulated in a lipid-dependent manner and modulates systemic fat uptake via ubiquitin ligase-dependent effects. Whether this metabolic regulation contributes to premature Parkinsonism warrants investigation.
Figures
Comment in
-
Parkin reinvents itself to regulate fatty acid metabolism by tagging CD36.J Clin Invest. 2011 Sep;121(9):3389-92. doi: 10.1172/JCI59219. Epub 2011 Aug 25. J Clin Invest. 2011. PMID: 21865651 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
