

Cost of adaptation and fitness effects of beneficial mutations in Pseudomonas fluorescens
- PMID: 21868607
- PMCID: PMC3213353
- DOI: 10.1534/genetics.111.130468
Cost of adaptation and fitness effects of beneficial mutations in Pseudomonas fluorescens
Abstract
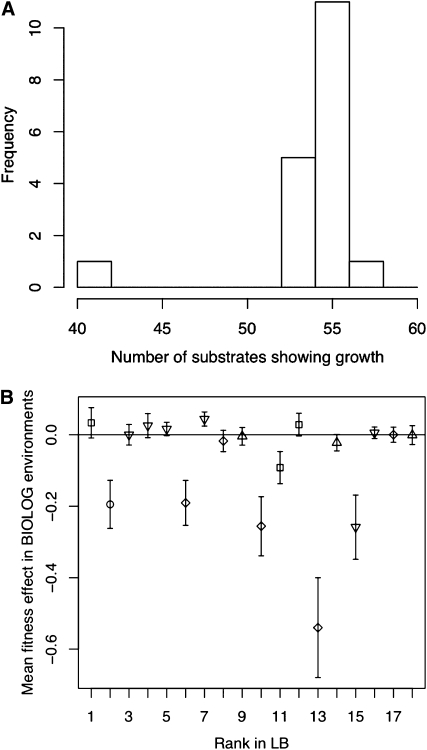
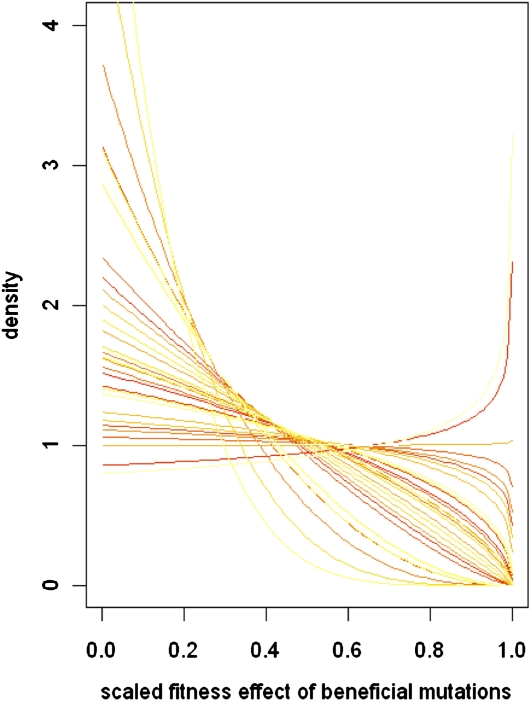
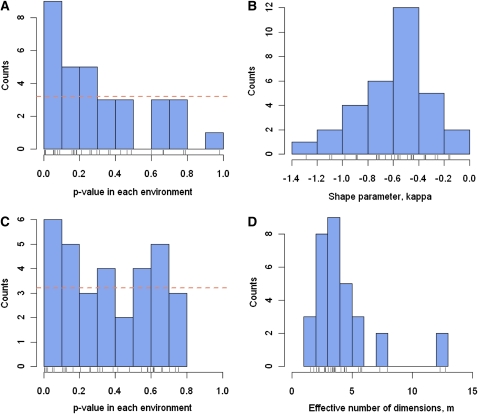
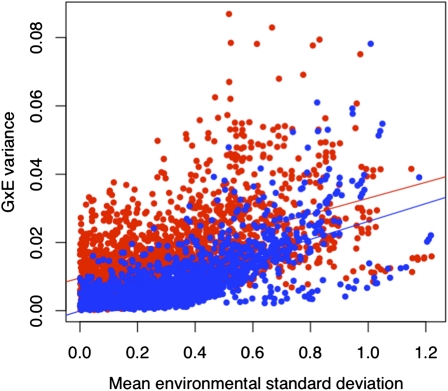
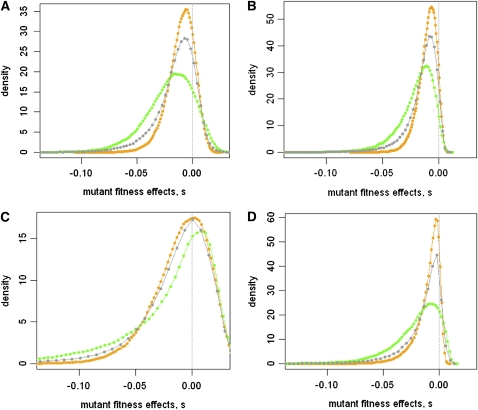
Adaptations are constructed through the sequential substitution of beneficial mutations by natural selection. However, the rarity of beneficial mutations has precluded efforts to describe even their most basic properties. Do beneficial mutations typically confer small or large fitness gains? Are their fitness effects environment specific, or are they broadly beneficial across a range of environments? To answer these questions, we used two subsets (n = 18 and n = 63) of a large library of mutants carrying antibiotic resistance mutations in the bacterium Pseudomonas fluorescens whose fitness, along with the antibiotic sensitive ancestor, was assayed across 95 novel environments differing in the carbon source available for growth. We explore patterns of genotype-by-environment (G × E) interactions and ecological specialization among the 18 mutants initially found superior to the sensitive ancestor in one environment. We find that G × E is remarkably similar between the two sets of mutants and that beneficial mutants are not typically associated with large costs of adaptation. Fitness effects among beneficial mutants depart from a strict exponential distribution: they assume a variety of shapes that are often roughly L shaped but always right truncated. Distributions of (beneficial) fitness effects predicted by a landscape model assuming multiple traits underlying fitness and a single optimum often provide a good description of the empirical distributions in our data. Simulations of data sets containing a mixture of single and double mutants under this landscape show that inferences about the distribution of fitness effects of beneficial mutants is quite robust to contamination by second-site mutations.
Figures
References
-
- Bell G., 1997. Selection: The Mechanism of Evolution. Chapman & Hall, London/New York
-
- Eyre-Walker A., Keightley P. D., 2007. The distribution of fitness effects of new mutations. Nat. Rev. Genet. 8: 610–618 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
