

What determines the activity of antimicrobial and cytolytic peptides in model membranes
- PMID: 21870782
- PMCID: PMC3200312
- DOI: 10.1021/bi200873u
What determines the activity of antimicrobial and cytolytic peptides in model membranes
Abstract
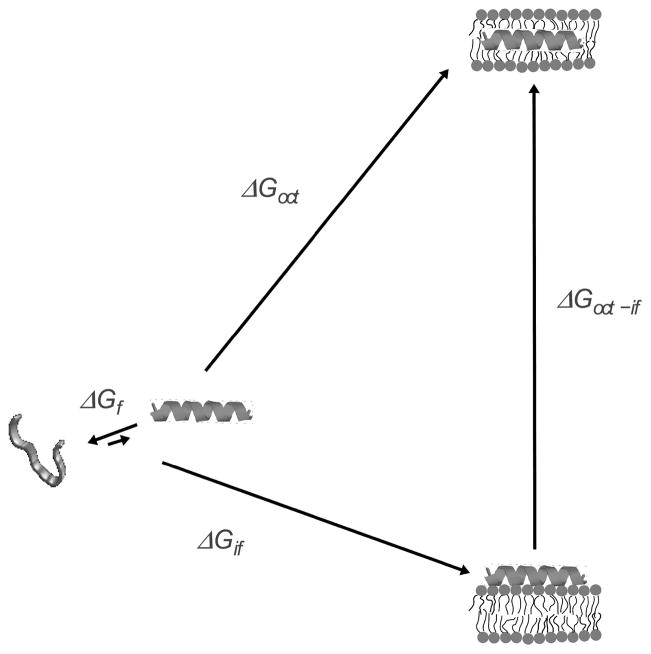
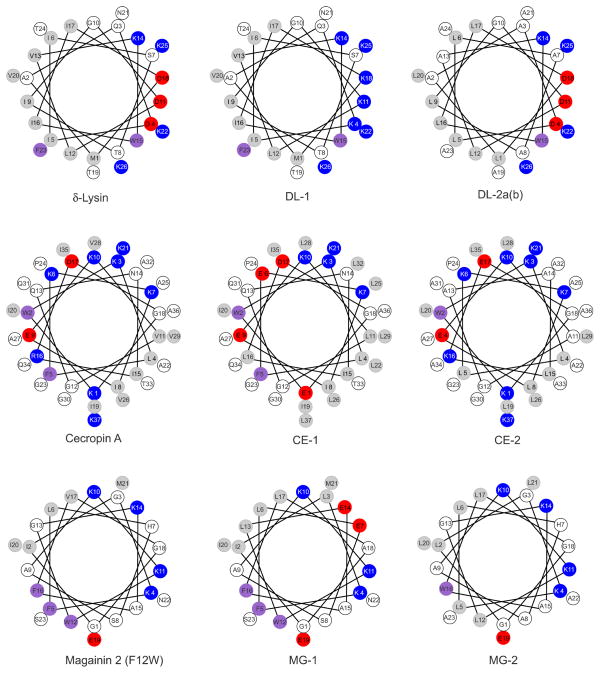
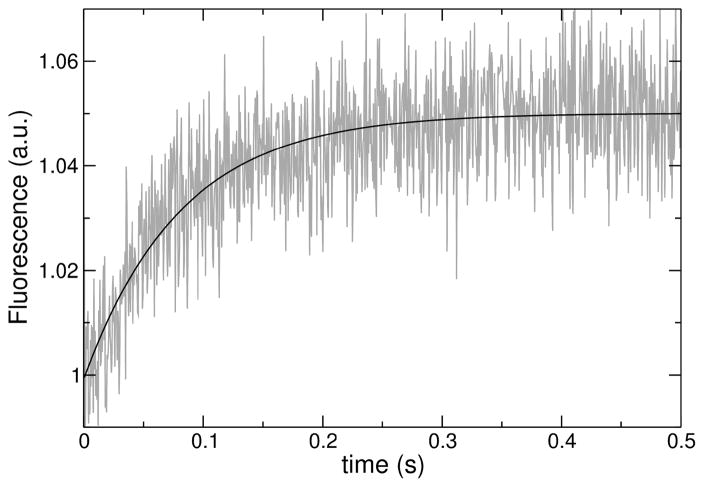
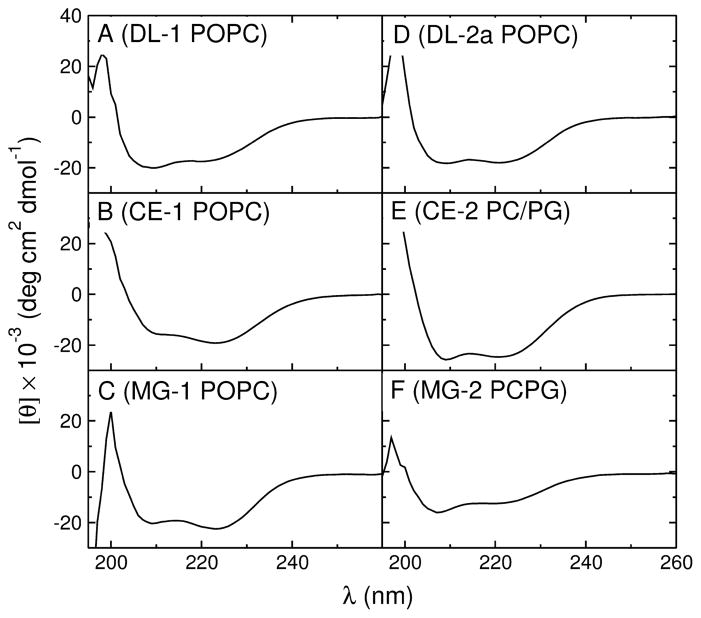
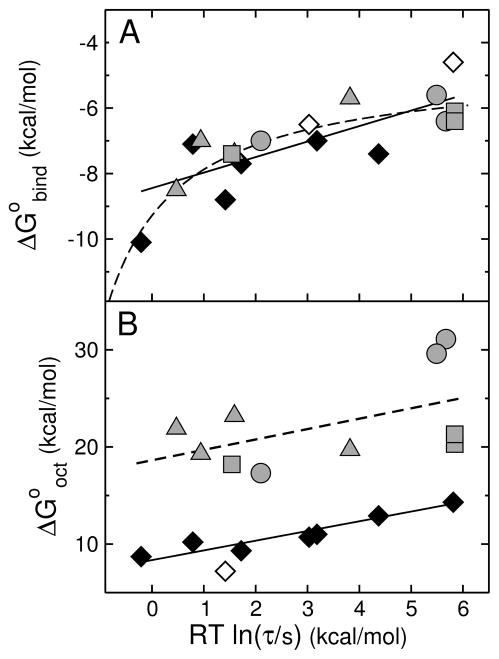
We previously proposed three hypotheses relating the mechanism of antimicrobial and cytolytic peptides in model membranes to the Gibbs free energies of binding and insertion into the membrane [Almeida, P. F., and Pokorny, A. (2009) Biochemistry 48, 8083-8093]. Two sets of peptides were designed to test those hypotheses, by mutating of the sequences of δ-lysin, cecropin A, and magainin 2. Peptide binding and activity were measured on phosphatidylcholine membranes. In the first set, the peptide charge was changed by mutating basic to acidic residues or vice versa, but the amino acid sequence was not altered much otherwise. The type of dye release changed from graded to all-or-none according to prediction. However, location of charged residues in the sequence with the correct spacing to form salt bridges failed to improve binding. In the second set, the charged and other key residues were kept in the same positions, whereas most of the sequence was significantly but conservatively simplified, maintaining the same hydrophobicity and amphipathicity. This set behaved completely different from predicted. The type of release, which was expected to be maintained, changed dramatically from all-or-none to graded in the mutants of cecropin and magainin. Finally, contrary to the hypotheses, the results indicate that the Gibbs energy of binding to the membrane, not the Gibbs energy of insertion, is the primary determinant of peptide activity.
© 2011 American Chemical Society
Figures
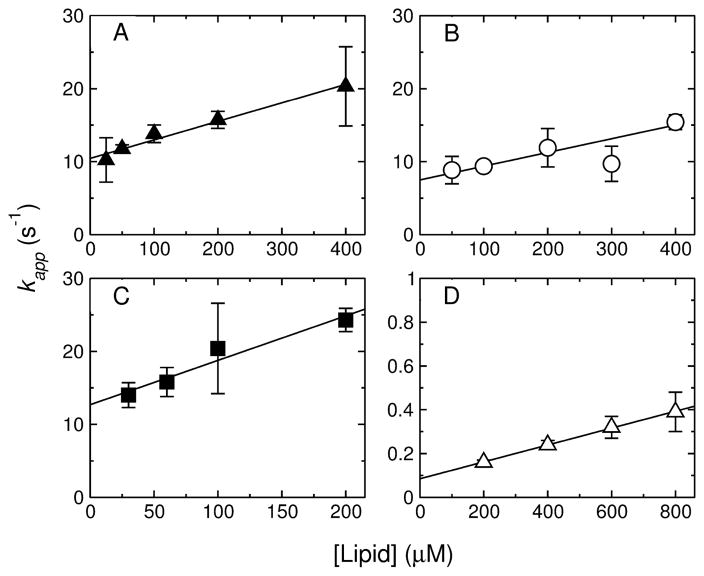
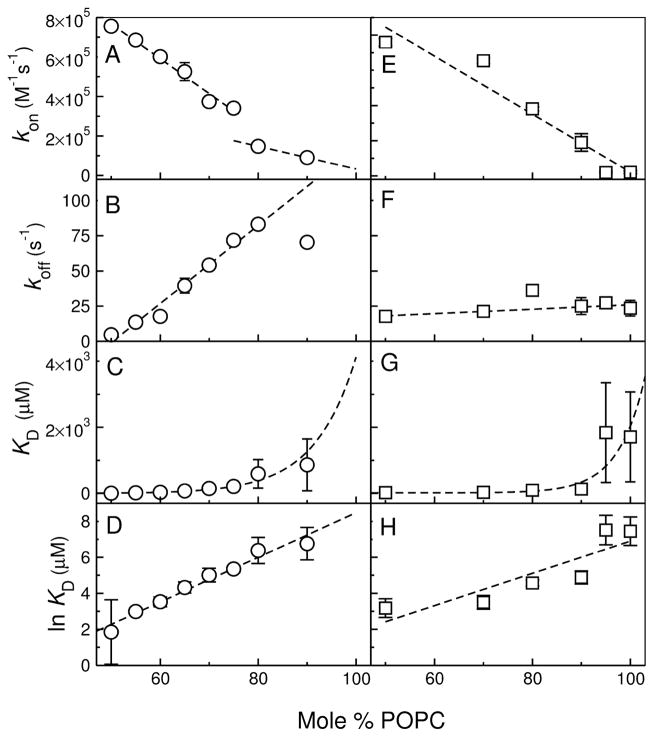
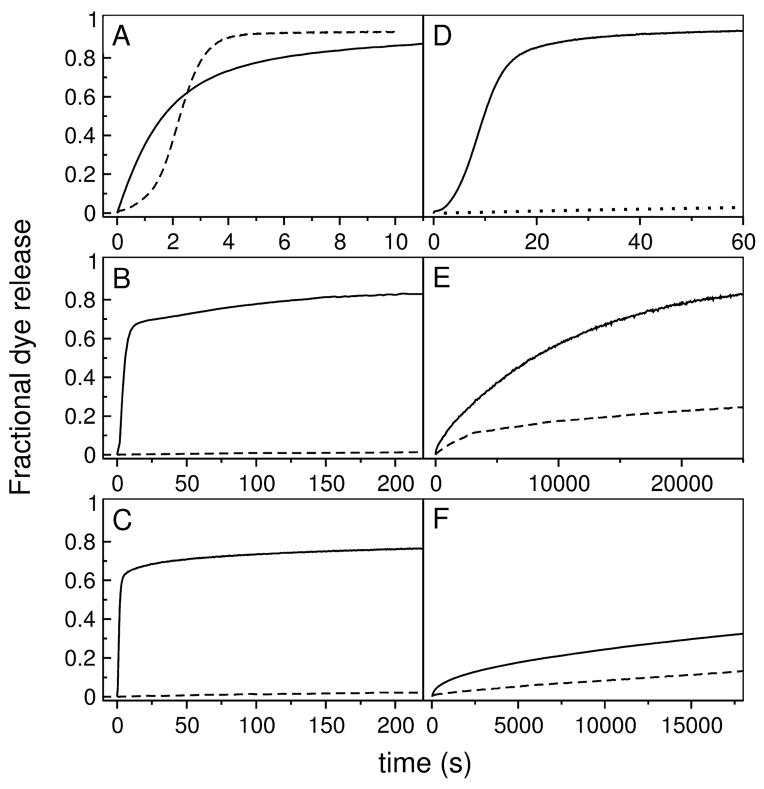
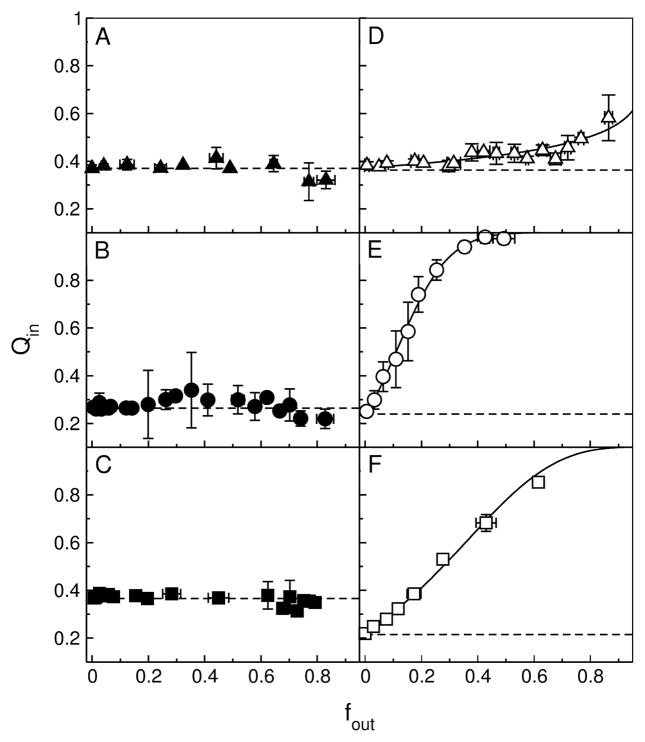
References
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389–395. - PubMed
-
- Giangaspero A, Sandri L, Tossi A. Amphipathic α-helical antimicrobial peptides. Eur J Biochem. 2001;268:5589–5600. - PubMed
-
- Tossi A, Sandri L, Giangaspero A. Amphipathic, α-helical antimicrobial peptides. Biopolymers. 2000;55:4–30. - PubMed
-
- Nickel W. The mystery of nonclassical protein secretion. A current view on cargo proteinsand potential export routes. Eur J Biochem. 2003;270:2109–2119. - PubMed
-
- Lindgren M, Langel U. Clasees and prediction of cell-penetrating peptides. Methods Mol Biol. 2011;683:3–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
