

Ten-Eleven-Translocation 2 (TET2) negatively regulates homeostasis and differentiation of hematopoietic stem cells in mice
- PMID: 21873190
- PMCID: PMC3167529
- DOI: 10.1073/pnas.1112317108
Ten-Eleven-Translocation 2 (TET2) negatively regulates homeostasis and differentiation of hematopoietic stem cells in mice
Abstract
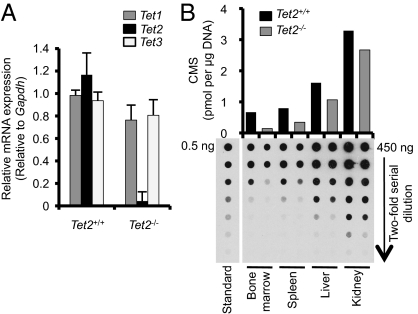
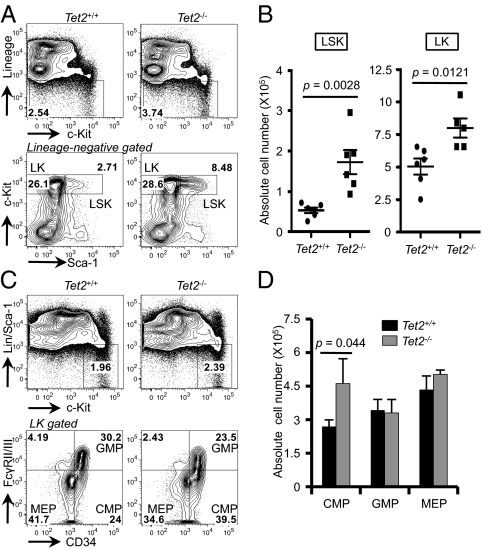
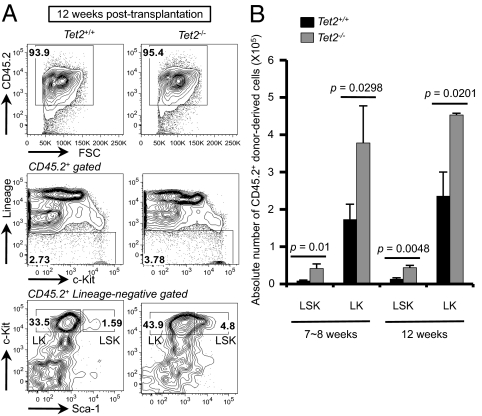
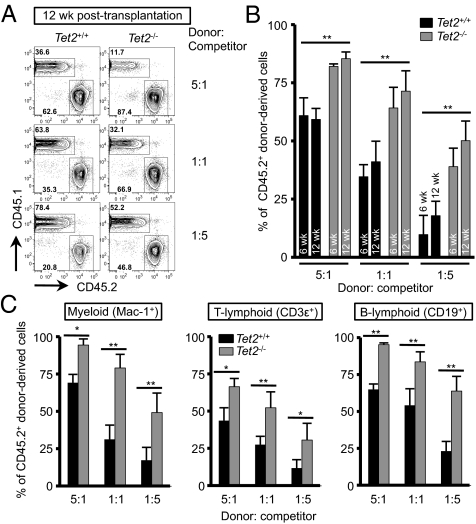
The Ten-Eleven-Translocation 2 (TET2) gene encodes a member of TET family enzymes that alters the epigenetic status of DNA by oxidizing 5-methylcytosine to 5-hydroxymethylcytosine (5hmC). Somatic loss-of-function mutations of TET2 are frequently observed in patients with diverse myeloid malignancies, including myelodysplastic syndromes, myeloproliferative neoplasms, and chronic myelomonocytic leukemia. By analyzing mice with targeted disruption of the Tet2 catalytic domain, we show here that Tet2 is a critical regulator of self-renewal and differentiation of hematopoietic stem cells (HSCs). Tet2 deficiency led to decreased genomic levels of 5hmC and augmented the size of the hematopoietic stem/progenitor cell pool in a cell-autonomous manner. In competitive transplantation assays, Tet2-deficient HSCs were capable of multilineage reconstitution and possessed a competitive advantage over wild-type HSCs, resulting in enhanced hematopoiesis into both lymphoid and myeloid lineages. In vitro, Tet2 deficiency delayed HSC differentiation and skewed development toward the monocyte/macrophage lineage. Our data indicate that Tet2 has a critical role in regulating the expansion and function of HSCs, presumably by controlling 5hmC levels at genes important for the self-renewal, proliferation, and differentiation of HSCs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
TET2 is essential for survival and hematopoietic stem cell homeostasis.Leukemia. 2012 Oct;26(10):2216-23. doi: 10.1038/leu.2012.94. Epub 2012 Apr 3. Leukemia. 2012. PMID: 22469782
-
Non-catalytic Roles of Tet2 Are Essential to Regulate Hematopoietic Stem and Progenitor Cell Homeostasis.Cell Rep. 2019 Sep 3;28(10):2480-2490.e4. doi: 10.1016/j.celrep.2019.07.094. Cell Rep. 2019. PMID: 31484061 Free PMC article.
-
Tet2 disruption leads to enhanced self-renewal and altered differentiation of fetal liver hematopoietic stem cells.Sci Rep. 2012;2:273. doi: 10.1038/srep00273. Epub 2012 Feb 17. Sci Rep. 2012. PMID: 22355785 Free PMC article.
-
TET2 as an epigenetic master regulator for normal and malignant hematopoiesis.Cancer Sci. 2014 Sep;105(9):1093-9. doi: 10.1111/cas.12484. Epub 2014 Sep 3. Cancer Sci. 2014. PMID: 25040794 Free PMC article. Review.
-
TET proteins and 5-methylcytosine oxidation in hematological cancers.Immunol Rev. 2015 Jan;263(1):6-21. doi: 10.1111/imr.12239. Immunol Rev. 2015. PMID: 25510268 Free PMC article. Review.
Cited by
-
Epigenetic control of myeloid cell differentiation, identity and function.Nat Rev Immunol. 2015 Jan;15(1):7-17. doi: 10.1038/nri3777. Nat Rev Immunol. 2015. PMID: 25534619 Review.
-
Genome-wide DNA hydroxymethylation changes are associated with neurodevelopmental genes in the developing human cerebellum.Hum Mol Genet. 2012 Dec 15;21(26):5500-10. doi: 10.1093/hmg/dds394. Epub 2012 Oct 5. Hum Mol Genet. 2012. PMID: 23042784 Free PMC article.
-
DNA Methylation in T-Cell Development and Differentiation.Crit Rev Immunol. 2020;40(2):135-156. doi: 10.1615/CritRevImmunol.2020033728. Crit Rev Immunol. 2020. PMID: 32749092 Free PMC article. Review.
-
TET2 coactivates gene expression through demethylation of enhancers.Sci Adv. 2018 Nov 7;4(11):eaau6986. doi: 10.1126/sciadv.aau6986. eCollection 2018 Nov. Sci Adv. 2018. PMID: 30417100 Free PMC article.
-
Epigenetic regulation of hematopoietic stem cell homeostasis.Blood Sci. 2019 Sep 17;1(1):19-28. doi: 10.1097/BS9.0000000000000018. eCollection 2019 Aug. Blood Sci. 2019. PMID: 35402787 Free PMC article. Review.
References
-
- Gal-Yam EN, Saito Y, Egger G, Jones PA. Cancer epigenetics: Modifications, screening, and therapy. Annu Rev Med. 2008;59:267–280. - PubMed
-
- Esteller M. Epigenetics in cancer. N Engl J Med. 2008;358:1148–1159. - PubMed
-
- Eden A, Gaudet F, Waghmare A, Jaenisch R. Chromosomal instability and tumors promoted by DNA hypomethylation. Science. 2003;300:455. - PubMed
-
- Gaudet F, et al. Induction of tumors in mice by genomic hypomethylation. Science. 2003;300:489–492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
