

PI3K/Akt signaling requires spatial compartmentalization in plasma membrane microdomains
- PMID: 21873248
- PMCID: PMC3167518
- DOI: 10.1073/pnas.1019386108
PI3K/Akt signaling requires spatial compartmentalization in plasma membrane microdomains
Abstract
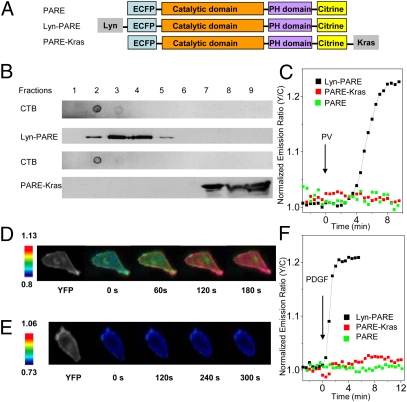
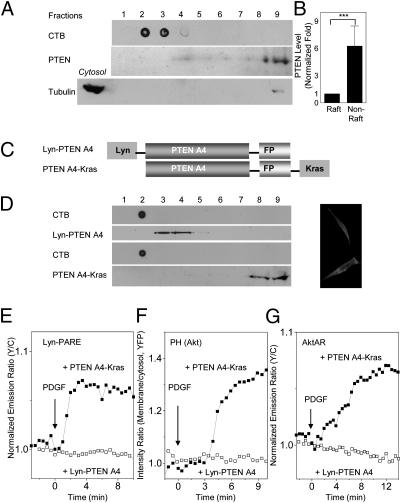
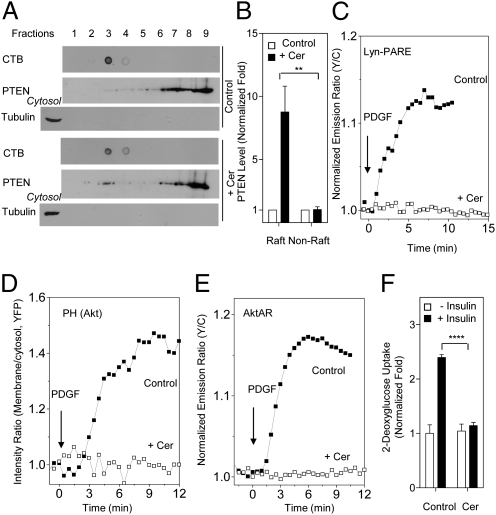
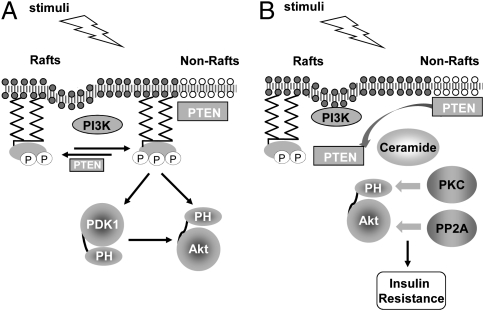
Spatial compartmentalization of signaling pathway components generally defines the specificity and enhances the efficiency of signal transduction. The phosphatidylinositol 3-kinase (PI3K)/Akt pathway is known to be compartmentalized within plasma membrane microdomains; however, the underlying mechanisms and functional impact of this compartmentalization are not well understood. Here, we show that phosphoinositide-dependent kinase 1 is activated in membrane rafts in response to growth factors, whereas the negative regulator of the pathway, phosphatase and tensin homolog deleted on chromosome 10 (PTEN), is primarily localized in nonraft regions. Alteration of this compartmentalization, either by genetic targeting or ceramide-induced recruitment of PTEN to rafts, abolishes the activity of the entire pathway. These findings reveal critical steps in raft-mediated PI3K/Akt activation and demonstrate the essential role of membrane microdomain compartmentalization in enabling PI3K/Akt signaling. They further suggest that dysregulation of this compartmentalization may underlie pathological complications such as insulin resistance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Franke TF. PI3K/Akt: Getting it right matters. Oncogene. 2008;27:6473–6488. - PubMed
-
- Farese RV, Sajan MP, Standaert ML. Insulin-sensitive protein kinases (atypical protein kinase C and protein kinase B/Akt): Actions and defects in obesity and type II diabetes. Exp Biol Med (Maywood) 2005;230:593–605. - PubMed
-
- Bozulic L, Hemmings BA. PIKKing on PKB: Regulation of PKB activity by phosphorylation. Curr Opin Cell Biol. 2009;21:256–261. - PubMed
-
- Mora A, Komander D, van Aalten DM, Alessi DR. PDK1, the master regulator of AGC kinase signal transduction. Semin Cell Dev Biol. 2004;15:161–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
