

Functional selectivity at the μ-opioid receptor: implications for understanding opioid analgesia and tolerance
- PMID: 21873412
- PMCID: PMC3186080
- DOI: 10.1124/pr.111.004598
Functional selectivity at the μ-opioid receptor: implications for understanding opioid analgesia and tolerance
Abstract
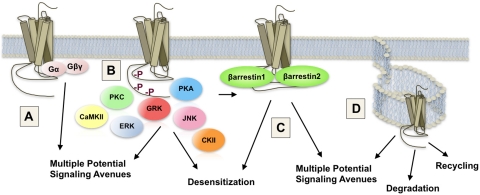
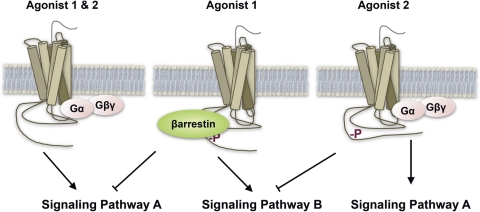
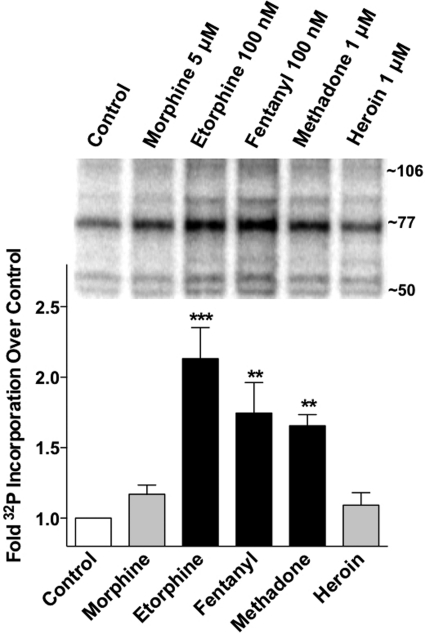
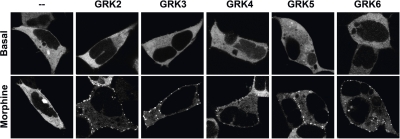
Opioids are the most effective analgesic drugs for the management of moderate or severe pain, yet their clinical use is often limited because of the onset of adverse side effects. Drugs in this class produce most of their physiological effects through activation of the μ opioid receptor; however, an increasing number of studies demonstrate that different opioids, while presumably acting at this single receptor, can activate distinct downstream responses, a phenomenon termed functional selectivity. Functional selectivity of receptor-mediated events can manifest as a function of the drug used, the cellular or neuronal environment examined, or the signaling or behavioral measure recorded. This review summarizes both in vitro and in vivo work demonstrating functional selectivity at the μ opioid receptor in terms of G protein coupling, receptor phosphorylation, interactions with β-arrestins, receptor desensitization, internalization and signaling, and details on how these differences may relate to the progression of analgesic tolerance after their extended use.
Figures
Similar articles
-
Effector antagonism by the regulators of G protein signalling (RGS) proteins causes desensitization of mu-opioid receptors in the CNS.Psychopharmacology (Berl). 2005 Jun;180(1):1-11. doi: 10.1007/s00213-005-2248-9. Epub 2005 Apr 14. Psychopharmacology (Berl). 2005. PMID: 15830230 Review.
-
β-arrestins: regulatory role and therapeutic potential in opioid and cannabinoid receptor-mediated analgesia.Handb Exp Pharmacol. 2014;219:427-43. doi: 10.1007/978-3-642-41199-1_22. Handb Exp Pharmacol. 2014. PMID: 24292843 Free PMC article. Review.
-
[Development of opioid tolerance -- molecular mechanisms and clinical consequences].Anasthesiol Intensivmed Notfallmed Schmerzther. 2003 Jan;38(1):14-26. doi: 10.1055/s-2003-36558. Anasthesiol Intensivmed Notfallmed Schmerzther. 2003. PMID: 12522725 Review. German.
-
Phosphorylation-deficient G-protein-biased μ-opioid receptors improve analgesia and diminish tolerance but worsen opioid side effects.Nat Commun. 2019 Jan 21;10(1):367. doi: 10.1038/s41467-018-08162-1. Nat Commun. 2019. PMID: 30664663 Free PMC article.
-
β-Arrestin-2 knockout prevents development of cellular μ-opioid receptor tolerance but does not affect opioid-withdrawal-related adaptations in single PAG neurons.Br J Pharmacol. 2015 Jan;172(2):492-500. doi: 10.1111/bph.12673. Epub 2014 Jul 1. Br J Pharmacol. 2015. PMID: 24597632 Free PMC article.
Cited by
-
Consequences of the 118A>G polymorphism in the OPRM1 gene: translation from bench to bedside?J Pain Res. 2013 May 1;6:331-53. doi: 10.2147/JPR.S42040. Print 2013. J Pain Res. 2013. PMID: 23658496 Free PMC article.
-
Functional selectivity of 6'-guanidinonaltrindole (6'-GNTI) at κ-opioid receptors in striatal neurons.J Biol Chem. 2013 Aug 2;288(31):22387-98. doi: 10.1074/jbc.M113.476234. Epub 2013 Jun 17. J Biol Chem. 2013. PMID: 23775075 Free PMC article.
-
Label-free integrative pharmacology on-target of opioid ligands at the opioid receptor family.BMC Pharmacol Toxicol. 2013 Mar 12;14:17. doi: 10.1186/2050-6511-14-17. BMC Pharmacol Toxicol. 2013. PMID: 23497702 Free PMC article.
-
Coordinated activity of a central pathway drives associative opioid analgesic tolerance.Sci Adv. 2023 Feb 10;9(6):eabo5627. doi: 10.1126/sciadv.abo5627. Epub 2023 Feb 8. Sci Adv. 2023. PMID: 36753548 Free PMC article.
-
Opioid signaling and design of analgesics.Prog Mol Biol Transl Sci. 2023;195:153-176. doi: 10.1016/bs.pmbts.2022.06.017. Epub 2022 Aug 5. Prog Mol Biol Transl Sci. 2023. PMID: 36707153 Free PMC article. Review.
References
-
- Abbadie C, Gultekin SH, Pasternak GW. (2000a) Immunohistochemical localization of the carboxy terminus of the novel mu opioid receptor splice variant MOR-1C within the human spinal cord. Neuroreport 11:1953–1957 - PubMed
-
- Abbadie C, Pan Y, Drake CT, Pasternak GW. (2000b) Comparative immunohistochemical distributions of carboxy terminus epitopes from the mu-opioid receptor splice variants MOR-1D, MOR-1 and MOR-1C in the mouse and rat CNS. Neuroscience 100:141–153 - PubMed
-
- Abbadie C, Pan YX, Pasternak GW. (2000c) Differential distribution in rat brain of mu opioid receptor carboxy terminal splice variants MOR-1C-like and MOR-1-like immunoreactivity: evidence for region-specific processing. J Comp Neurol 419:244–256 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
