

Enhanced Ca2+ entry due to Orai1 plasma membrane insertion increases IL-8 secretion by cystic fibrosis airways
- PMID: 21873556
- PMCID: PMC3236623
- DOI: 10.1096/fj.11-187682
Enhanced Ca2+ entry due to Orai1 plasma membrane insertion increases IL-8 secretion by cystic fibrosis airways
Abstract
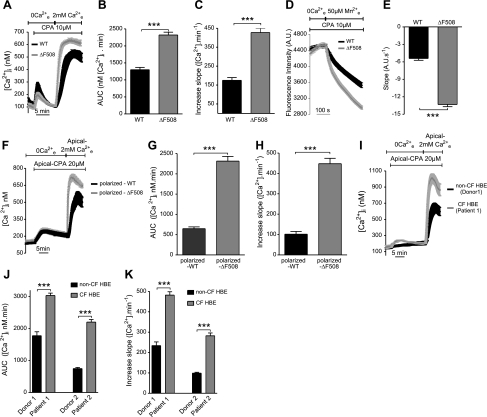
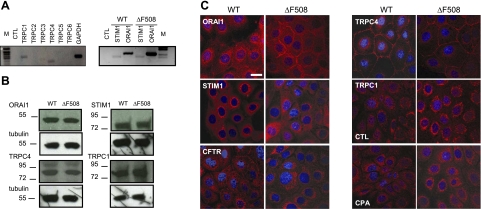
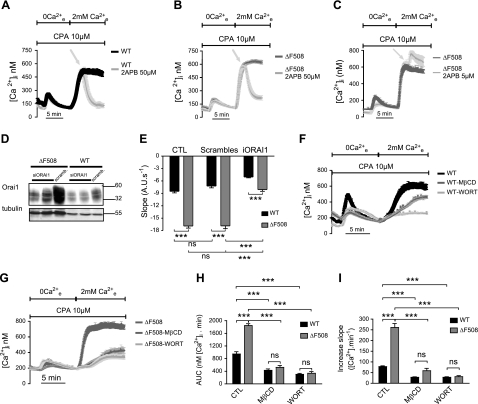
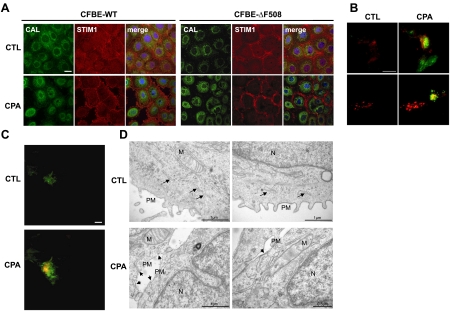
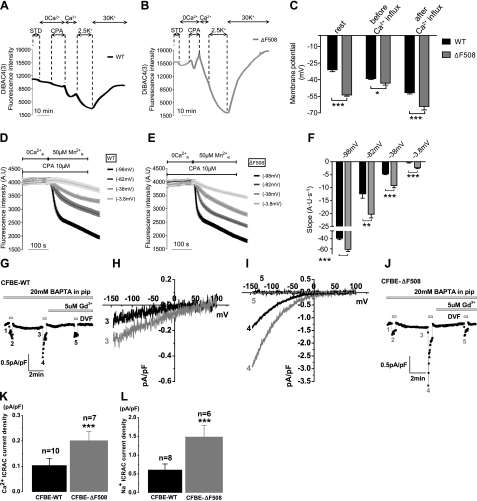
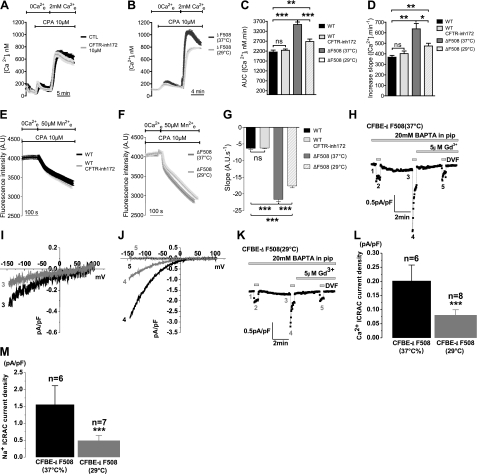
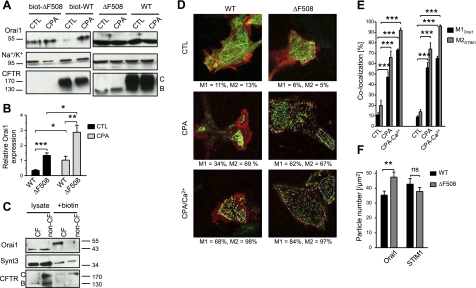
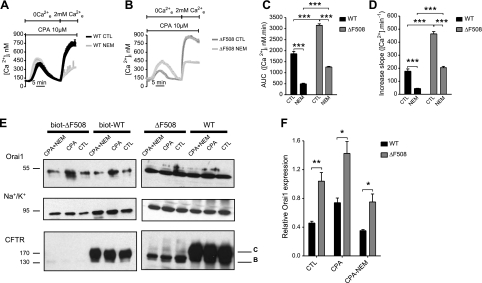
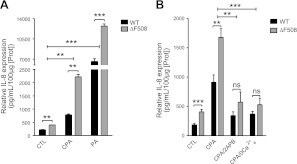
Cystic fibrosis (CF) is caused by mutations in the gene encoding the CF transmembrane conductance regulator (CFTR). The most common mutation, ΔF508, causes retention of CFTR in the endoplasmic reticulum (ER). Some CF abnormalities can be explained by altered Ca(2+) homeostasis, although it remains unknown how CFTR influences calcium signaling. This study examined the novel hypothesis that store-operated calcium entry (SOCE) through Orai1 is abnormal in CF. The significance of Orai1-mediated SOCE for increased interleukin-8 (IL-8) expression in CF was also investigated. CF and non-CF human airway epithelial cell line and primary cells (obtained at lung transplantation) were used in Ca(2+) imaging, electrophysiology, and fluorescence imaging experiments to explore differences in Orai1 function in CF vs. non-CF cells. Protein expression and localization was assessed by Western blots, cell surface biotinylation, ELISA, and image correlation spectroscopy (ICS). We show here that store-operated Ca(2+) entry (SOCE) is elevated in CF human airway epithelial cells (hAECs; ≈ 1.8- and ≈ 2.5-fold for total Ca(2+)(i) increase and Ca(2+) influx rate, respectively, and ≈ 2-fold increase in the I(CRAC) current) and is caused by increased exocytotic insertion (≈ 2-fold) of Orai1 channels into the plasma membrane, which is normalized by rescue of ΔF508-CFTR trafficking to the cell surface. Augmented SOCE in CF cells is a major factor leading to increased IL-8 secretion (≈ 2-fold). CFTR normally down-regulates the Orai1/stromal interaction molecule 1 (STIM1) complex, and loss of this inhibition due to the absence of CFTR at the plasma membrane helps to explain the potentiated inflammatory response in CF cells.
Figures
References
-
- Rommens J. M., Zengerling S., Burns J., Melmer G., Kerem B. S., Plavsic N., Zsiga M., Kennedy D., Markiewicz D., Rozmahel R., Riordan J. R., Buchwald M., Tsui L.-C. (1988) Identification and regional localization of DNA markers on chromosome 7 for the cloning of the cystic fibrosis gene. Am. J. Hum. Genet. 43, 645–663 - PMC - PubMed
-
- Riordan J. R., Rommens J. M., Kerem B., Alon N., Rozmahel R., Grzelczak Z., Zielenski J., Lok S., Plavsic N., Chou J. L., Drumm M. L., Iannuzzi M. C., Collins F. S., Tsui L. C. (1989) Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245, 1066–1073 - PubMed
-
- Hanrahan J. W., Wioland M. A. (2004) Revisiting cystic fibrosis transmembrane conductance regulator structure and function. Proc. Am. Thorac. Soc. 1, 17–21 - PubMed
-
- Naren A. P., Cobb B., Li C., Roy K., Nelson D., Heda G. D., Liao J., Kirk K. L., Sorscher E. J., Hanrahan J., Clancy J. P. (2003) A macromolecular complex of beta 2 adrenergic receptor, CFTR, and ezrin/radixin/moesin-binding phosphoprotein 50 is regulated by PKA. Proc. Natl. Acad. Sci. U. S. A. 100, 342–346 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
