

Mechanism of actin filament nucleation by Vibrio VopL and implications for tandem W domain nucleation
- PMID: 21873985
- PMCID: PMC3173040
- DOI: 10.1038/nsmb.2109
Mechanism of actin filament nucleation by Vibrio VopL and implications for tandem W domain nucleation
Abstract
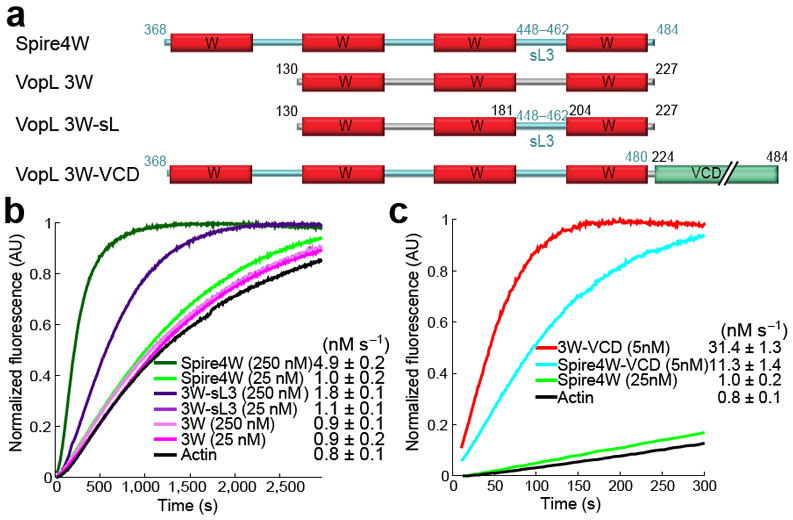
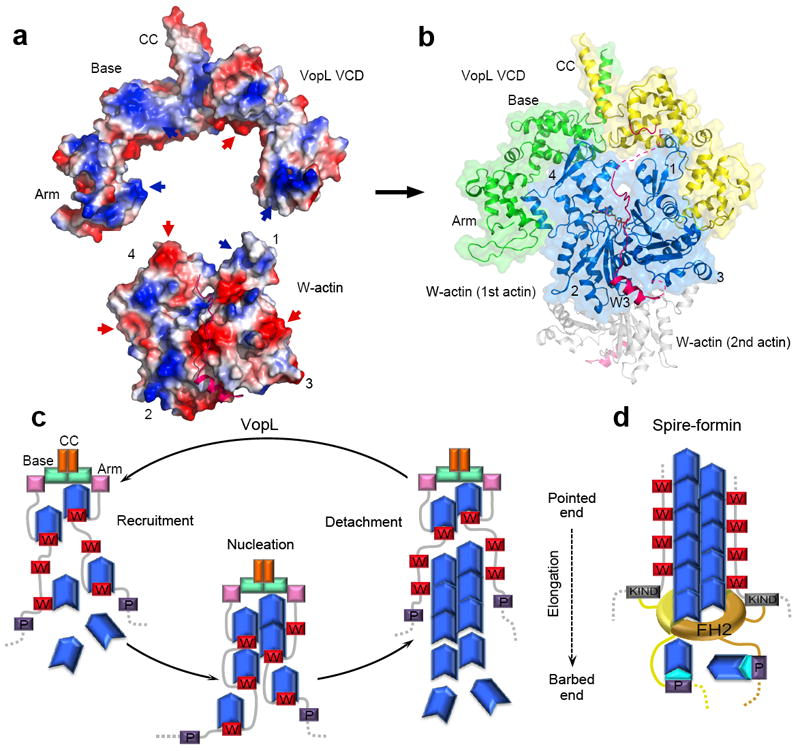
Pathogen proteins targeting the actin cytoskeleton often serve as model systems to understand their more complex eukaryotic analogs. We show that the strong actin filament nucleation activity of Vibrio parahaemolyticus VopL depends on its three W domains and on its dimerization through a unique VopL C-terminal domain (VCD). The VCD shows a previously unknown all-helical fold and interacts with the pointed end of the actin nucleus, contributing to the nucleation activity directly and through duplication of the W domain repeat. VopL promotes rapid cycles of filament nucleation and detachment but generally has no effect on elongation. Profilin inhibits VopL-induced nucleation by competing for actin binding to the W domains. Combined, the results suggest that VopL stabilizes a hexameric double-stranded pointed end nucleus. Analysis of hybrid constructs of VopL and the eukaryotic nucleator Spire suggest that Spire may also function as a dimer in cells.
Figures




Comment in
-
A new twist in actin filament nucleation.Nat Struct Mol Biol. 2011 Sep 6;18(9):967-9. doi: 10.1038/nsmb.2130. Nat Struct Mol Biol. 2011. PMID: 21894217 No abstract available.
References
-
- Pollard TD. Regulation of actin filament assembly by Arp2/3 complex and formins. Annu Rev Biophys Biomol Struct. 2007;36:451–77. - PubMed
-
- Goode BL, Eck MJ. Mechanism and function of formins in the control of actin assembly. Annu Rev Biochem. 2007;76:593–627. - PubMed
-
- Higgs HN. Formin proteins: a domain-based approach. Trends Biochem Sci. 2005;30:342–53. - PubMed
-
- Quinlan ME, Heuser JE, Kerkhoff E, Mullins RD. Drosophila Spire is an actin nucleation factor. Nature. 2005;433:382–8. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
