

Drosophila Rae1 controls the abundance of the ubiquitin ligase Highwire in post-mitotic neurons
- PMID: 21874015
- PMCID: PMC3183334
- DOI: 10.1038/nn.2922
Drosophila Rae1 controls the abundance of the ubiquitin ligase Highwire in post-mitotic neurons
Abstract
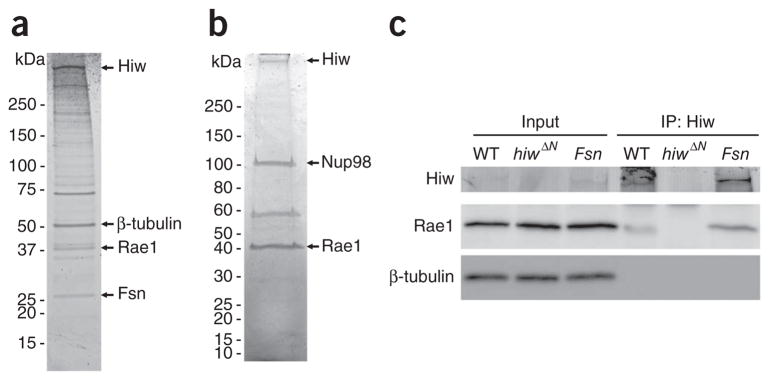
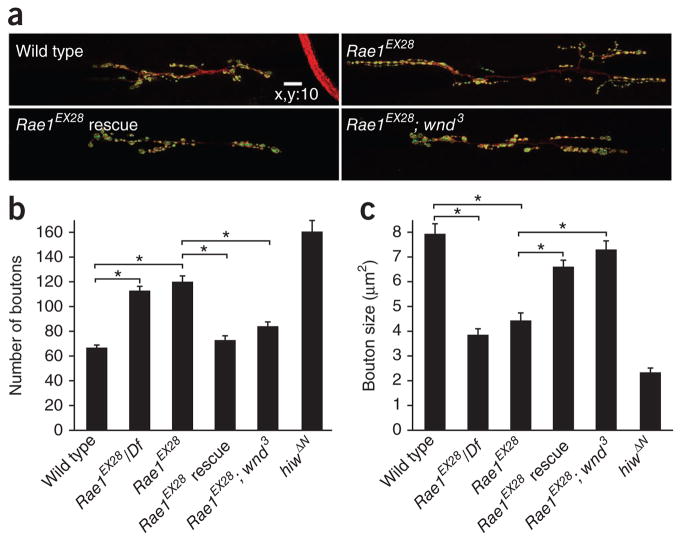
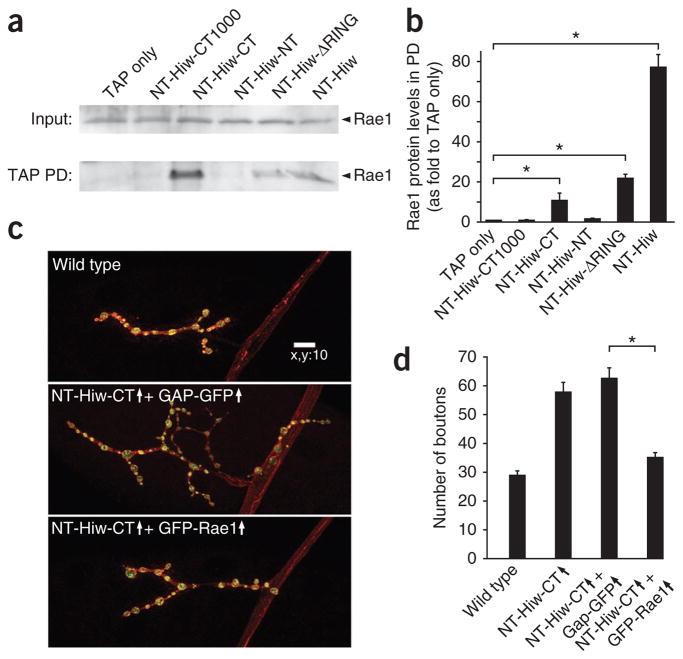
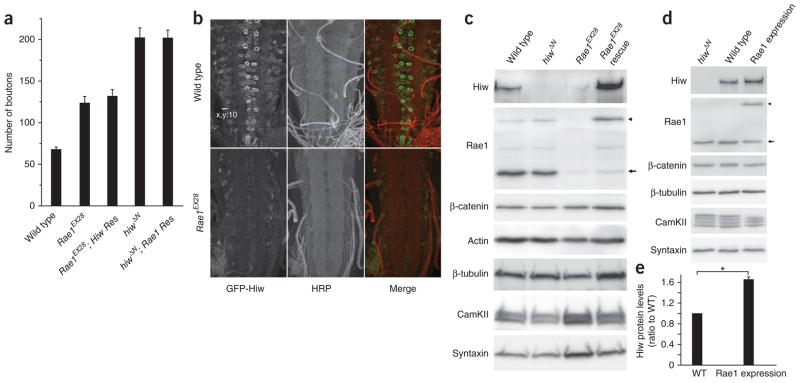
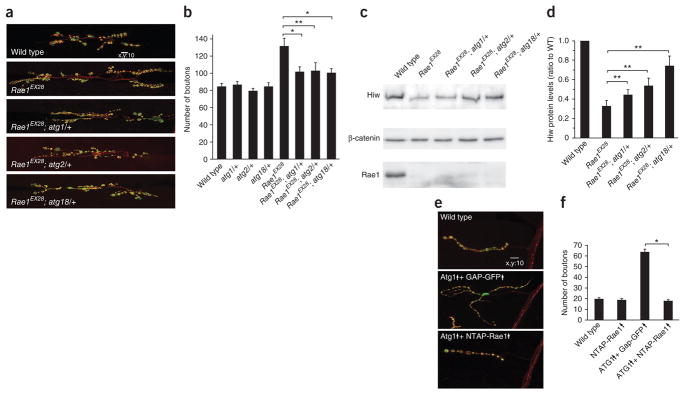
The evolutionarily conserved Highwire (Hiw)/Drosophila Fsn E3 ubiquitin ligase complex is required for normal synaptic morphology during development and axonal regeneration after injury. However, little is known about the molecular mechanisms that regulate the Hiw E3 ligase complex. Using tandem affinity purification techniques, we identified Drosophila Rae1 as a previously unknown component of the Hiw/Fsn complex. Loss of Rae1 function in neurons results in morphological defects at the neuromuscular junction that are similar to those seen in hiw mutants. We found that Rae1 physically and genetically interacts with Hiw and restrains synaptic terminal growth by regulating the MAP kinase kinase kinase Wallenda. Moreover, we found that the Rae1 is both necessary and sufficient to promote Hiw protein abundance, and it does so by binding to Hiw and protecting Hiw from autophagy-mediated degradation. These results describe a previously unknown mechanism that selectively controls Hiw protein abundance during synaptic development.
© 2011 Nature America, Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



Comment in
-
Maintaining a Highwire act.Nat Neurosci. 2011 Sep 27;14(10):1226. doi: 10.1038/nn1011-1226. Nat Neurosci. 2011. PMID: 21952265 No abstract available.
References
-
- Po MD, Hwang C, Zhen M. PHRs: bridging axon guidance, outgrowth and synapse development. Curr Opin Neurobiol. 2010;20:100–107. - PubMed
-
- DiAntonio A, et al. Ubiquitination-dependent mechanisms regulate synaptic growth and function. Nature. 2001;412:449–452. - PubMed
-
- Wan HI, et al. Highwire regulates synaptic growth in Drosophila. Neuron. 2000;26:313–329. - PubMed
-
- Schaefer AM, Hadwiger GD, Nonet ML. rpm-1, a conserved neuronal gene that regulates targeting and synaptogenesis in C. elegans. Neuron. 2000;26:345–356. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
