

A widespread class of reverse transcriptase-related cellular genes
- PMID: 21876125
- PMCID: PMC3251080
- DOI: 10.1073/pnas.1100266108
A widespread class of reverse transcriptase-related cellular genes
Abstract
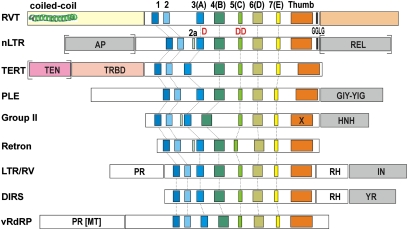
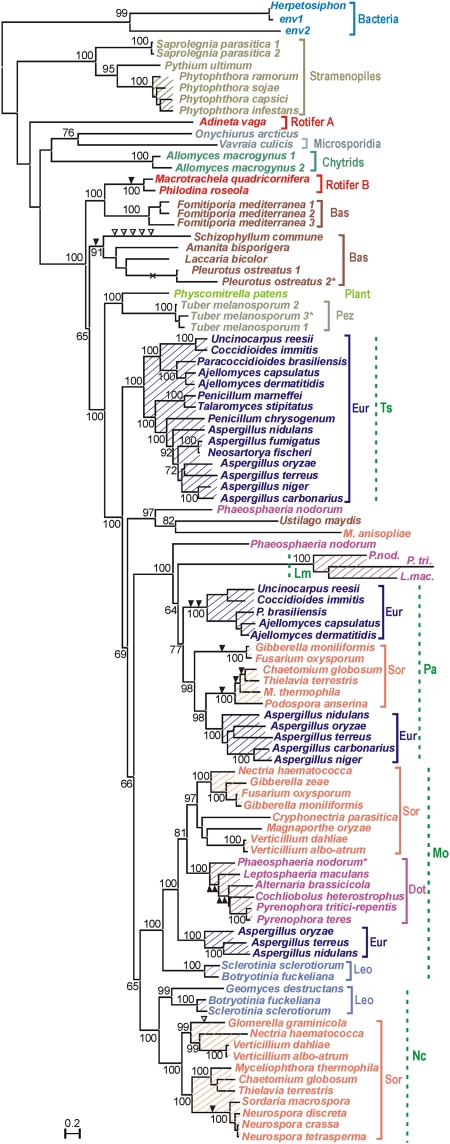
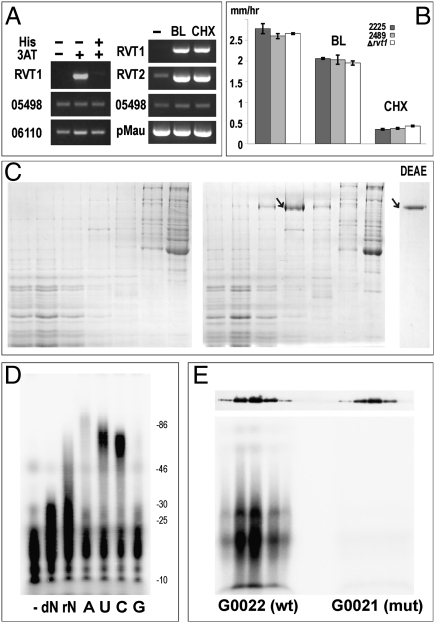
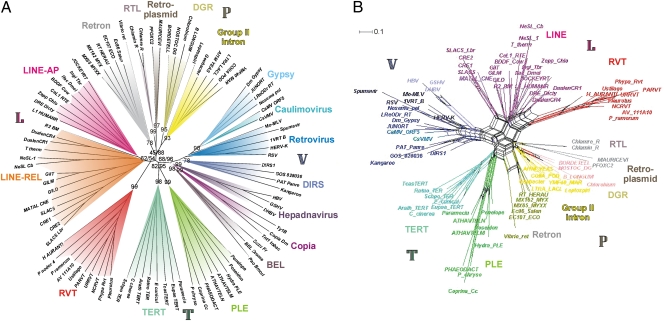
Reverse transcriptases (RTs) polymerize DNA on RNA templates. They fall into several structurally related but distinct classes and form an assemblage of RT-like enzymes that, in addition to RTs, also includes certain viral RNA-dependent RNA polymerases (RdRP) synthesizing RNA on RNA templates. It is generally believed that most RT-like enzymes originate from retrotransposons or viruses and have no specific function in the host cell, with telomerases being the only notable exception. Here we report on the discovery and properties of a unique class of RT-related cellular genes collectively named rvt. We present evidence that rvts are not components of retrotransposons or viruses, but single-copy genes with a characteristic domain structure that may contain introns in evolutionarily conserved positions, occur in syntenic regions, and evolve under purifying selection. These genes can be found in all major taxonomic groups including protists, fungi, animals, plants, and even bacteria, although they exhibit patchy phylogenetic distribution in each kingdom. We also show that the RVT protein purified from one of its natural hosts, Neurospora crassa, exists in a multimeric form and has the ability to polymerize NTPs as well as dNTPs in vitro, with a strong preference for NTPs, using Mn(2+) as a cofactor. The existence of a previously unknown class of single-copy RT-related genes calls for reevaluation of the current views on evolution and functional roles of RNA-dependent polymerases in living cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Crick F. Central dogma of molecular biology. Nature. 1970;227:561–563. - PubMed
-
- Baltimore D. RNA-dependent DNA polymerase in virions of RNA tumour viruses. Nature. 1970;226:1209–1211. - PubMed
-
- Temin HM, Mizutani S. RNA-dependent DNA polymerase in virions of Rous sarcoma virus. Nature. 1970;226:1211–1213. - PubMed
-
- Eickbush TH, Malik H. Origins and evolution of retrotransposons. In: Craig NL, Craigie R, Gellert M, Lambowitz AM, editors. Mobile DNA II. Washington, DC: ASM; 2002. pp. 1111–1144.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
