

Global discovery of small RNAs in Yersinia pseudotuberculosis identifies Yersinia-specific small, noncoding RNAs required for virulence
- PMID: 21876162
- PMCID: PMC3174644
- DOI: 10.1073/pnas.1101655108
Global discovery of small RNAs in Yersinia pseudotuberculosis identifies Yersinia-specific small, noncoding RNAs required for virulence
Abstract
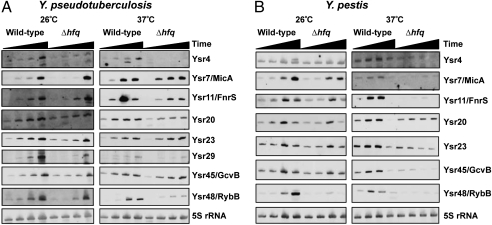
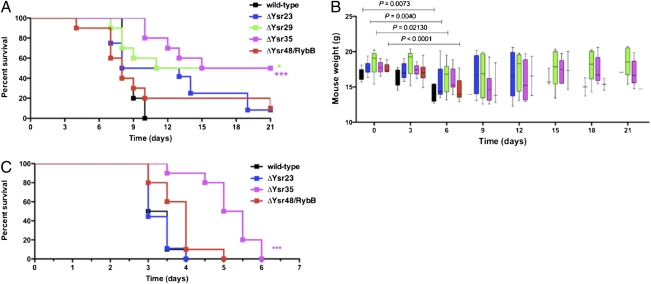
A major class of bacterial small, noncoding RNAs (sRNAs) acts by base-pairing with mRNAs to alter the translation from and/or stability of the transcript. Our laboratory has shown that Hfq, the chaperone that mediates the interaction of many sRNAs with their targets, is required for the virulence of the enteropathogen Yersinia pseudotuberculosis. This finding suggests that sRNAs play a critical role in the regulation of virulence in this pathogen, but these sRNAs are not known. Using a deep sequencing approach, we identified the global set of sRNAs expressed in vitro by Y. pseudotuberculosis. Sequencing of RNA libraries from bacteria grown at 26 °C and 37 °C resulted in the identification of 150 unannotated sRNAs. The majority of these sRNAs are Yersinia specific, without orthologs in either Escherichia coli or Salmonella typhimurium. Six sRNAs are Y. pseudotuberculosis specific and are absent from the genome of the closely related species Yersinia pestis. We found that the expression of many sRNAs conserved between Y. pseudotuberculosis and Y. pestis differs in both timing and dependence on Hfq, suggesting evolutionary changes in posttranscriptional regulation between these species. Deletion of multiple sRNAs in Y. pseudotuberculosis leads to attenuation of the pathogen in a mouse model of yersiniosis, as does the inactivation in Y. pestis of a conserved, Yersinia-specific sRNA in a mouse model of pneumonic plague. Finally, we determined the regulon controlled by one of these sRNAs, revealing potential virulence determinants in Y. pseudotuberculosis that are regulated in a posttranscriptional manner.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Sharma CM, Vogel J. Experimental approaches for the discovery and characterization of regulatory small RNA. Curr Opin Microbiol. 2009;12:536–546. - PubMed
-
- Gottesman S. The small RNA regulators of Escherichia coli: Roles and mechanisms. Annu Rev Microbiol. 2004;58:303–328. - PubMed
-
- Vogel J, Papenfort K. Small non-coding RNAs and the bacterial outer membrane. Curr Opin Microbiol. 2006;9:605–611. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
