

Conserved meiotic machinery in Glomus spp., a putatively ancient asexual fungal lineage
- PMID: 21876220
- PMCID: PMC3184777
- DOI: 10.1093/gbe/evr089
Conserved meiotic machinery in Glomus spp., a putatively ancient asexual fungal lineage
Abstract
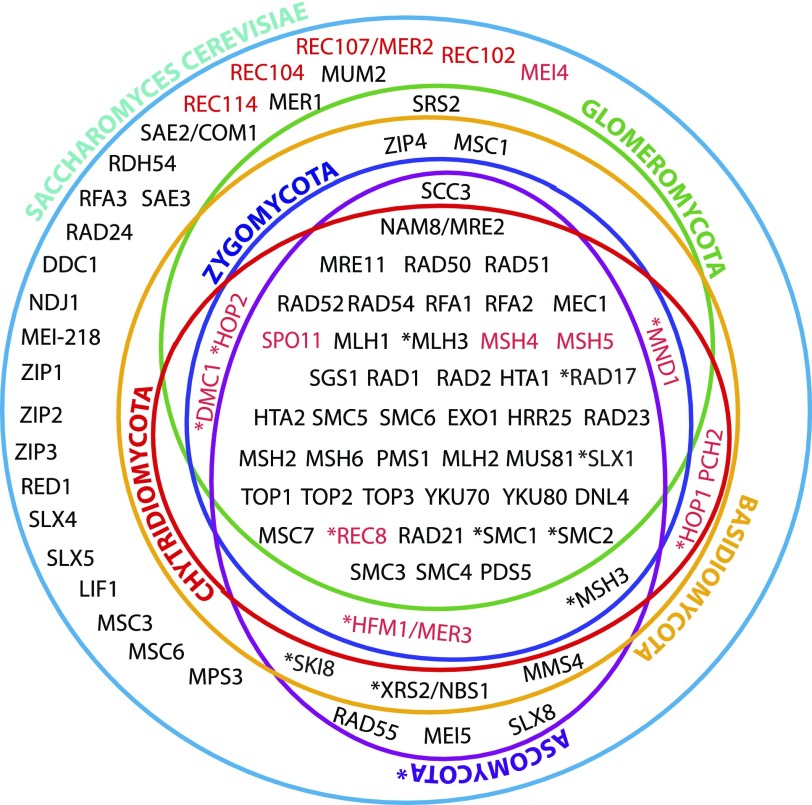
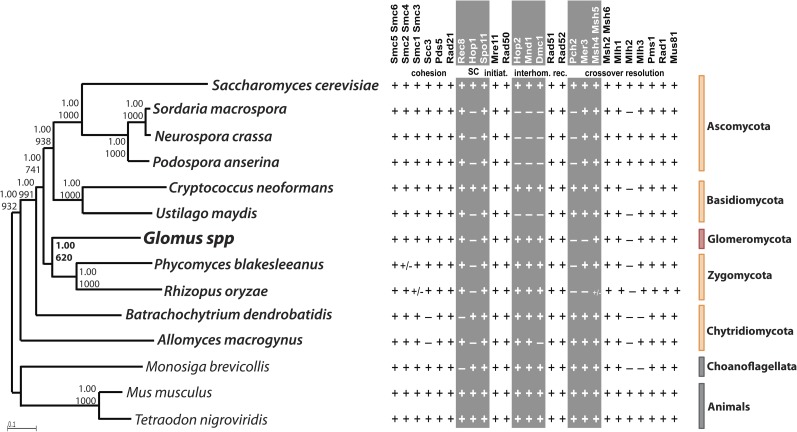
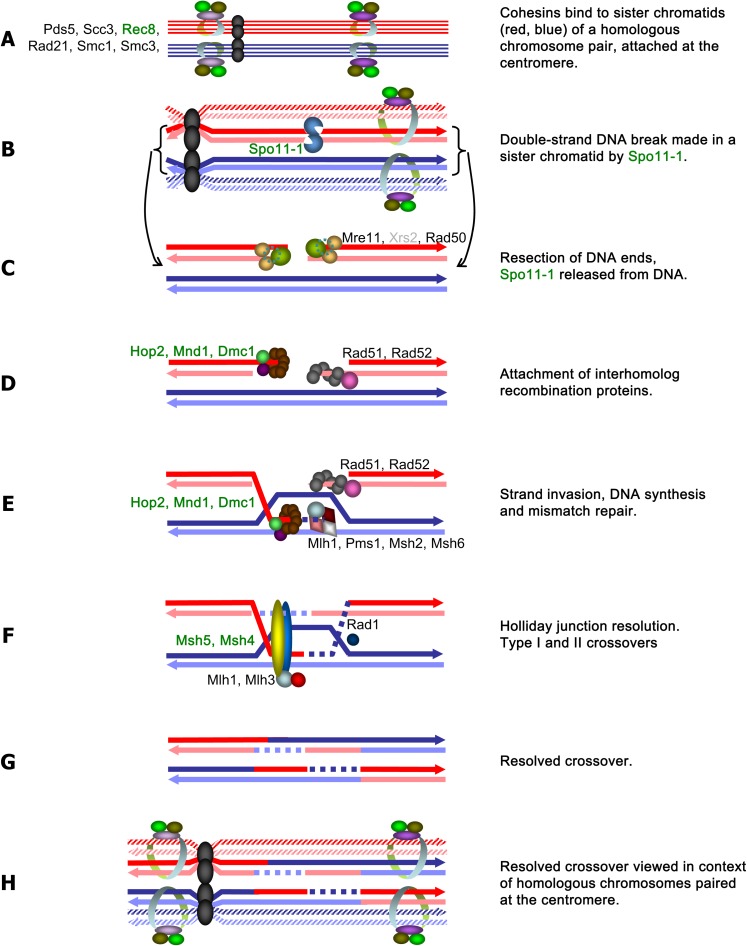
Arbuscular mycorrhizal fungi (AMF) represent an ecologically important and evolutionarily intriguing group of symbionts of land plants, currently thought to have propagated clonally for over 500 Myr. AMF produce multinucleate spores and may exchange nuclei through anastomosis, but meiosis has never been observed in this group. A provocative alternative for their successful and long asexual evolutionary history is that these organisms may have cryptic sex, allowing them to recombine alleles and compensate for deleterious mutations. This is partly supported by reports of recombination among some of their natural populations. We explored this hypothesis by searching for some of the primary tools for a sustainable sexual cycle--the genes whose products are required for proper completion of meiotic recombination in yeast--in the genomes of four AMF and compared them with homologs of representative ascomycete, basidiomycete, chytridiomycete, and zygomycete fungi. Our investigation used molecular and bioinformatic tools to identify homologs of 51 meiotic genes, including seven meiosis-specific genes and other "core meiotic genes" conserved in the genomes of the AMF Glomus diaphanum (MUCL 43196), Glomus irregulare (DAOM-197198), Glomus clarum (DAOM 234281), and Glomus cerebriforme (DAOM 227022). Homology of AMF meiosis-specific genes was verified by phylogenetic analyses with representative fungi, animals (Mus, Hydra), and a choanoflagellate (Monosiga). Together, these results indicate that these supposedly ancient asexual fungi may be capable of undergoing a conventional meiosis; a hypothesis that is consistent with previous reports of recombination within and across some of their populations.
Figures
Similar articles
-
Fungal sex: meiosis machinery in ancient symbiotic fungi.Curr Biol. 2011 Nov 8;21(21):R896-7. doi: 10.1016/j.cub.2011.09.021. Curr Biol. 2011. PMID: 22075432
-
Recombination in Glomus intraradices, a supposed ancient asexual arbuscular mycorrhizal fungus.BMC Evol Biol. 2009 Jan 15;9:13. doi: 10.1186/1471-2148-9-13. BMC Evol Biol. 2009. PMID: 19146661 Free PMC article.
-
Conserved Proteins of the RNA Interference System in the Arbuscular Mycorrhizal Fungus Rhizoglomus irregulare Provide New Insight into the Evolutionary History of Glomeromycota.Genome Biol Evol. 2018 Jan 1;10(1):328-343. doi: 10.1093/gbe/evy002. Genome Biol Evol. 2018. PMID: 29329439 Free PMC article.
-
Lineage-Specific Genes and Cryptic Sex: Parallels and Differences between Arbuscular Mycorrhizal Fungi and Fungal Pathogens.Trends Plant Sci. 2021 Feb;26(2):111-123. doi: 10.1016/j.tplants.2020.09.006. Epub 2020 Sep 30. Trends Plant Sci. 2021. PMID: 33011084 Review.
-
Using a meiosis detection toolkit to investigate ancient asexual "scandals" and the evolution of sex.Bioessays. 2008 Jun;30(6):579-89. doi: 10.1002/bies.20764. Bioessays. 2008. PMID: 18478537 Review.
Cited by
-
Rethinking asexuality: the enigmatic case of functional sexual genes in Lepraria (Stereocaulaceae).BMC Genomics. 2024 Oct 26;25(1):1003. doi: 10.1186/s12864-024-10898-8. BMC Genomics. 2024. PMID: 39455957 Free PMC article.
-
Basidioascus undulatus: genome, origins, and sexuality.IMA Fungus. 2015 Jun;6(1):215-31. doi: 10.5598/imafungus.2015.06.01.14. Epub 2015 Jun 19. IMA Fungus. 2015. PMID: 26203425 Free PMC article.
-
Recombination between clonal lineages of the asexual fungus Verticillium dahliae detected by genotyping by sequencing.PLoS One. 2014 Sep 2;9(9):e106740. doi: 10.1371/journal.pone.0106740. eCollection 2014. PLoS One. 2014. PMID: 25181515 Free PMC article.
-
Sex in cheese: evidence for sexuality in the fungus Penicillium roqueforti.PLoS One. 2012;7(11):e49665. doi: 10.1371/journal.pone.0049665. Epub 2012 Nov 21. PLoS One. 2012. PMID: 23185400 Free PMC article.
-
The frequency of sex in fungi.Philos Trans R Soc Lond B Biol Sci. 2016 Oct 19;371(1706):20150540. doi: 10.1098/rstb.2015.0540. Philos Trans R Soc Lond B Biol Sci. 2016. PMID: 27619703 Free PMC article. Review.
References
-
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005;21:2104–2105. - PubMed
-
- Ackerman S, Kermany AR, Hickey DA. Finite populations, finite resources, and the evolutionary maintenance of genetic recombination. J Hered. 2010;101:S135–S141. - PubMed
-
- Angelard C, Colard A, Niculita-Hirzel H, Croll D, Sanders IR. Segregation in a mycorrhizal fungus alters rice growth and symbiosis-specific gene transcription. Curr Biol. 2010;20:1216–1221. - PubMed
-
- Archetti M. Complementation, genetic conflict, and the evolution of sex and recombination. J Hered. 2010;101:S21–S33. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
