

Protective antiviral antibody responses in a mouse model of influenza virus infection require TACI
- PMID: 21881204
- PMCID: PMC3195469
- DOI: 10.1172/JCI57362
Protective antiviral antibody responses in a mouse model of influenza virus infection require TACI
Abstract
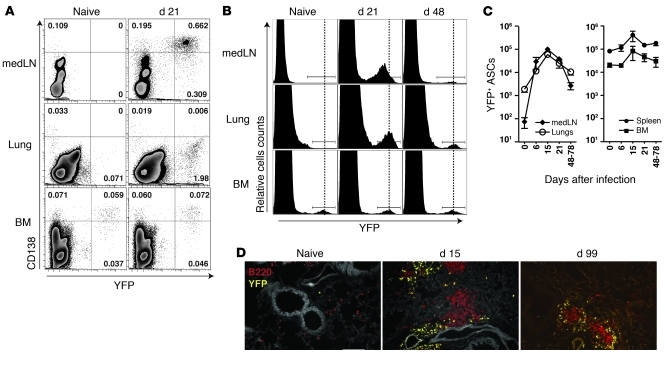
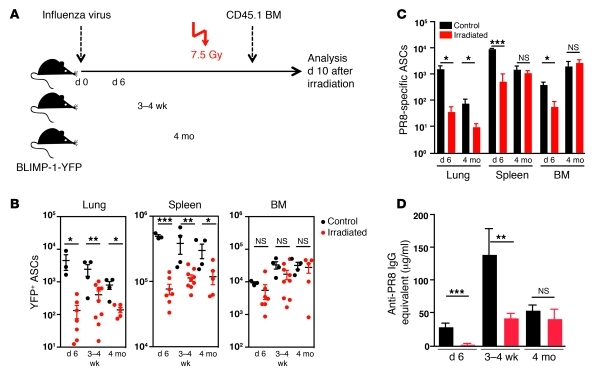
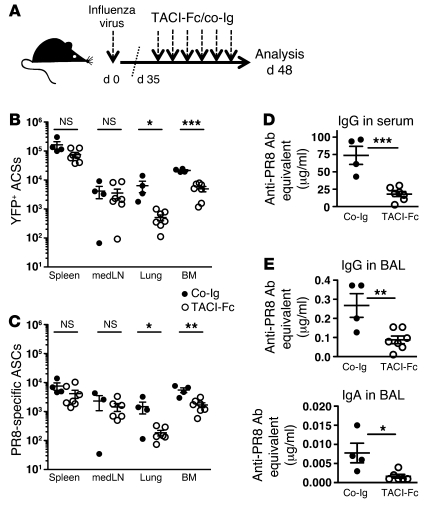
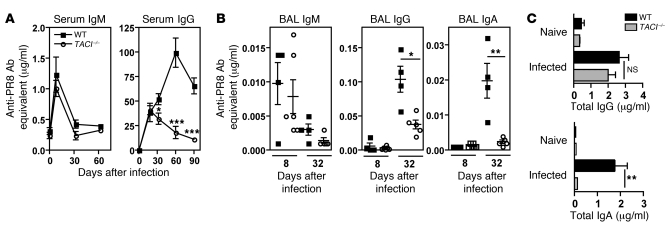
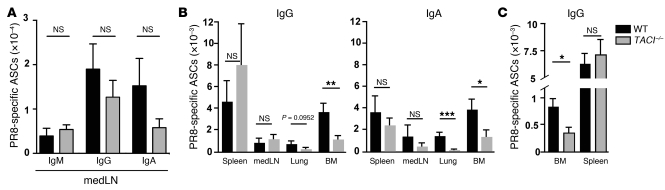
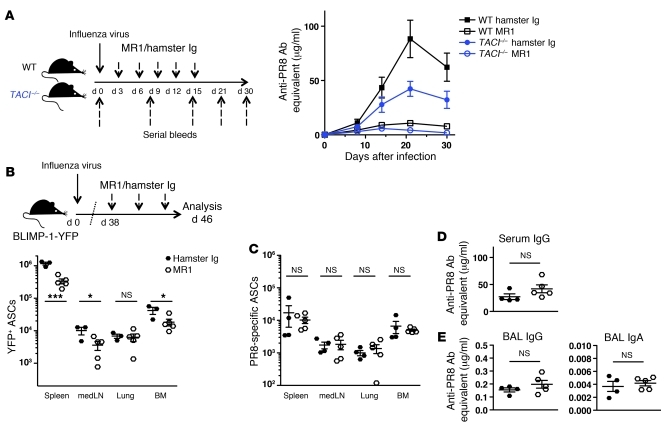
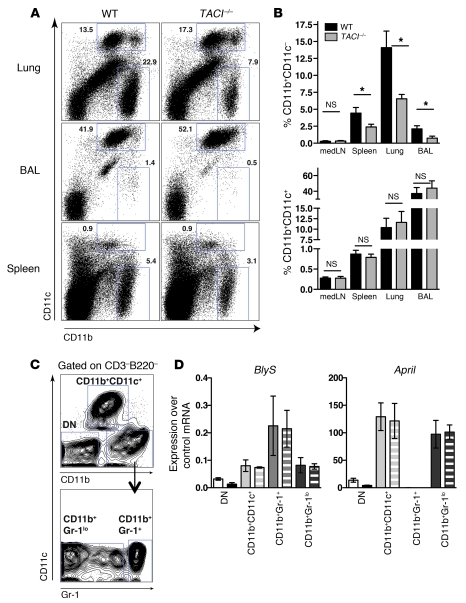
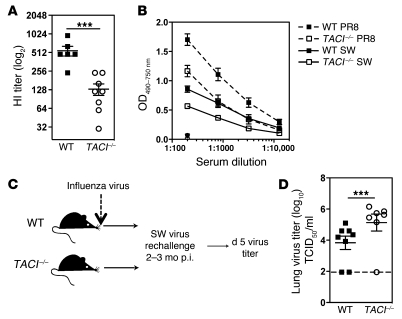
Antiviral Abs, for example those produced in response to influenza virus infection, are critical for virus neutralization and defense against secondary infection. While the half-life of Abs is short, Ab titers can last a lifetime due to a subset of the Ab-secreting cells (ASCs) that is long lived. However, the mechanisms governing ASC longevity are poorly understood. Here, we have identified a critical role for extrinsic cytokine signals in the survival of respiratory tract ASCs in a mouse model of influenza infection. Irradiation of mice at various time points after influenza virus infection markedly diminished numbers of lung ASCs, suggesting that they are short-lived and require extrinsic factors in order to persist. Neutralization of the TNF superfamily cytokines B lymphocyte stimulator (BLyS; also known as BAFF) and a proliferation-inducing ligand (APRIL) reduced numbers of antiviral ASCs in the lungs and bone marrow, whereas ASCs in the spleen and lung-draining lymph node were surprisingly unaffected. Mice deficient in transmembrane activator and calcium-modulator and cyclophilin ligand interactor (TACI), a receptor for BLyS and APRIL, mounted an initial antiviral B cell response similar to that generated in WT mice but failed to sustain protective Ab titers in the airways and serum, leading to increased susceptibility to secondary viral challenge. These studies highlight the importance of TACI signaling for the maintenance of ASCs and protection against influenza virus infection.
Figures
Similar articles
-
Deficient TACI expression on B lymphocytes of newborn mice leads to defective Ig secretion in response to BAFF or APRIL.J Immunol. 2008 Jul 15;181(2):976-90. doi: 10.4049/jimmunol.181.2.976. J Immunol. 2008. PMID: 18606649
-
TACI, unlike BAFF-R, is solely activated by oligomeric BAFF and APRIL to support survival of activated B cells and plasmablasts.Blood. 2008 Feb 1;111(3):1004-12. doi: 10.1182/blood-2007-09-110874. Epub 2007 Oct 17. Blood. 2008. PMID: 17942754
-
The role of the BAFF/APRIL system in the T cell-independent specific response to blood stage Plasmodium falciparum hemozoin.Cytokine. 2018 Nov;111:445-453. doi: 10.1016/j.cyto.2018.05.037. Epub 2018 Jun 5. Cytokine. 2018. PMID: 29884307
-
Transmembrane activator and calcium-modulator and cyclophilin ligand interactor mutations in common variable immunodeficiency.Curr Opin Allergy Clin Immunol. 2008 Dec;8(6):520-6. doi: 10.1097/ACI.0b013e3283141200. Curr Opin Allergy Clin Immunol. 2008. PMID: 18978466 Review.
-
The role of the BAFF/APRIL system on T cell function.Semin Immunol. 2006 Oct;18(5):284-9. doi: 10.1016/j.smim.2006.04.005. Epub 2006 Aug 22. Semin Immunol. 2006. PMID: 16931039 Review.
Cited by
-
Immunoregulation by antibody secreting cells in inflammation, infection, and cancer.Immunol Rev. 2021 Sep;303(1):103-118. doi: 10.1111/imr.12991. Epub 2021 Jun 17. Immunol Rev. 2021. PMID: 34145601 Free PMC article. Review.
-
Vaccines against respiratory viral pathogens for use in neonates: opportunities and challenges.J Immunol. 2014 Dec 1;193(11):5363-9. doi: 10.4049/jimmunol.1401410. J Immunol. 2014. PMID: 25411431 Free PMC article. Review.
-
Immune Cell Dynamics in Rhesus Macaques Infected with a Brazilian Strain of Zika Virus.J Immunol. 2017 Aug 1;199(3):1003-1011. doi: 10.4049/jimmunol.1700256. Epub 2017 Jun 30. J Immunol. 2017. PMID: 28667164 Free PMC article.
-
Type I Interferon Regulates the Survival and Functionality of B Cells in Rainbow Trout.Front Immunol. 2020 Jul 9;11:1494. doi: 10.3389/fimmu.2020.01494. eCollection 2020. Front Immunol. 2020. PMID: 32733485 Free PMC article.
-
Pulmonary infection with influenza A virus induces site-specific germinal center and T follicular helper cell responses.PLoS One. 2012;7(7):e40733. doi: 10.1371/journal.pone.0040733. Epub 2012 Jul 11. PLoS One. 2012. PMID: 22792401 Free PMC article.
References
-
- Gerhard W. The role of the antibody response in influenza virus infection. Curr Top Microbiol Immunol. 2001;260:171–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 AI055428/AI/NIAID NIH HHS/United States
- R01 AI071087/AI/NIAID NIH HHS/United States
- P30 CA010815/CA/NCI NIH HHS/United States
- P01 AI061093/AI/NIAID NIH HHS/United States
- P30 CA10815/CA/NCI NIH HHS/United States
- T32 CA009171/CA/NCI NIH HHS/United States
- AI52861/AI/NIAID NIH HHS/United States
- AI061093/AI/NIAID NIH HHS/United States
- U19 AI083022/AI/NIAID NIH HHS/United States
- R01 AI052861/AI/NIAID NIH HHS/United States
- AI071087/AI/NIAID NIH HHS/United States
- R01 AI054488/AI/NIAID NIH HHS/United States
- T32 CA09171-32/CA/NCI NIH HHS/United States
- AG58066/AG/NIA NIH HHS/United States
- R01 AI093722/AI/NIAID NIH HHS/United States
- AI055428/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
