

A mutation in the immunoproteasome subunit PSMB8 causes autoinflammation and lipodystrophy in humans
- PMID: 21881205
- PMCID: PMC3195477
- DOI: 10.1172/JCI58414
A mutation in the immunoproteasome subunit PSMB8 causes autoinflammation and lipodystrophy in humans
Abstract
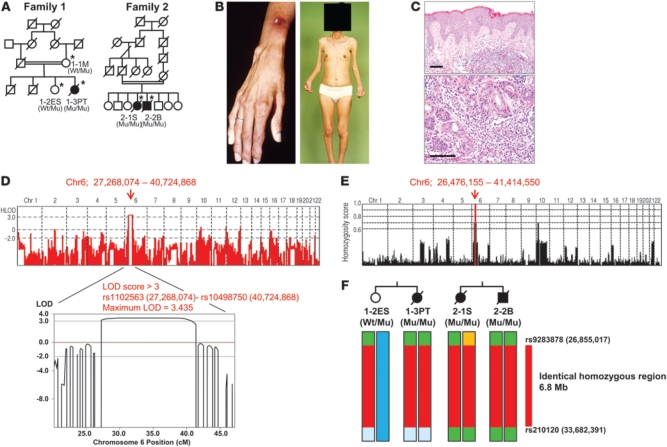
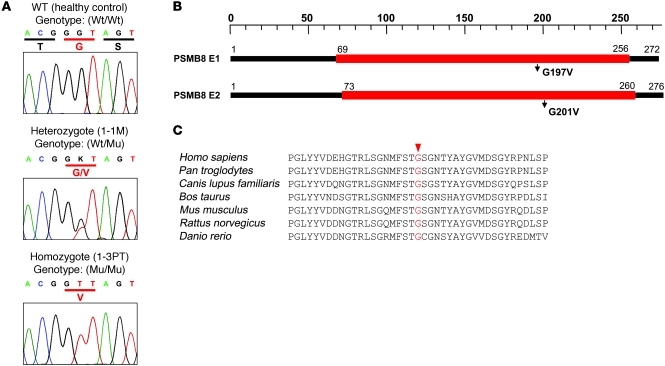
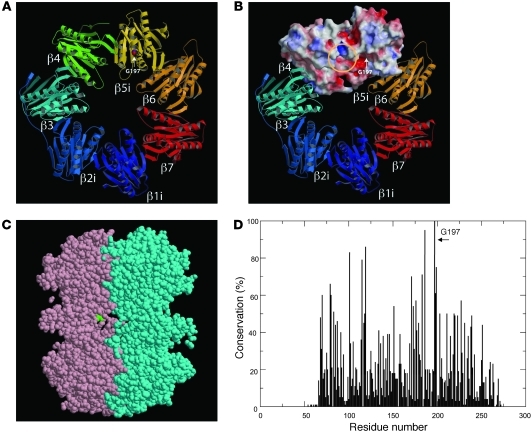
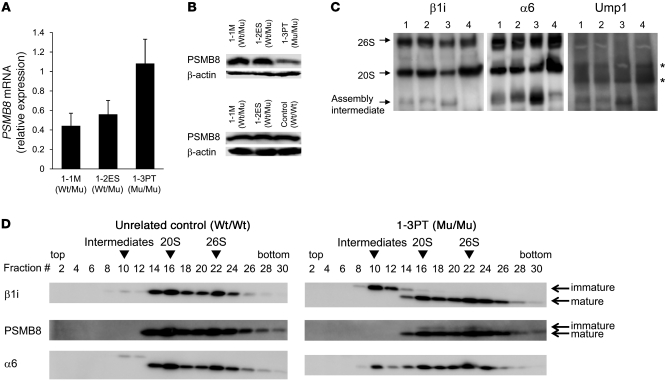
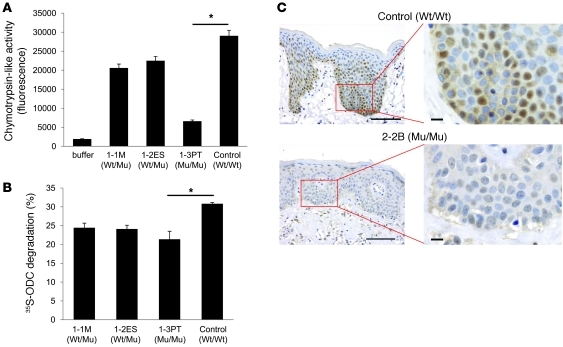
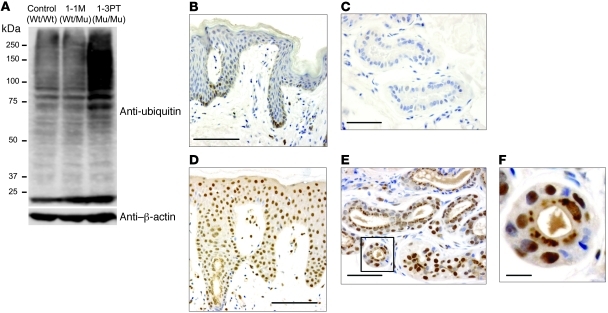
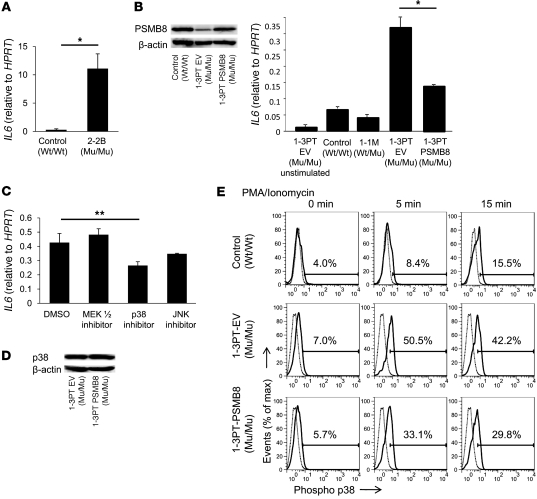
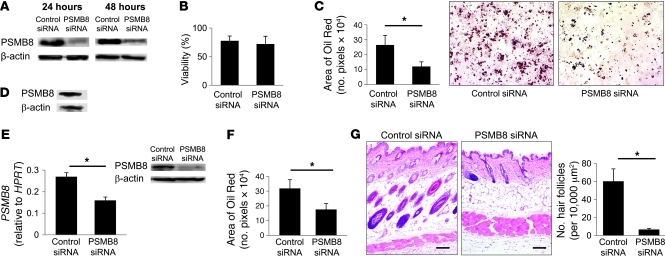
Proteasomes are multisubunit proteases that play a critical role in maintaining cellular function through the selective degradation of ubiquitinated proteins. When 3 additional β subunits, expression of which is induced by IFN-γ, are substituted for their constitutively expressed counterparts, the structure is converted to an immunoproteasome. However, the underlying roles of immunoproteasomes in human diseases are poorly understood. Using exome analysis, we found a homozygous missense mutation (G197V) in immunoproteasome subunit, β type 8 (PSMB8), which encodes one of the β subunits induced by IFN-γ in patients from 2 consanguineous families. Patients bearing this mutation suffered from autoinflammatory responses that included recurrent fever and nodular erythema together with lipodystrophy. This mutation increased assembly intermediates of immunoproteasomes, resulting in decreased proteasome function and ubiquitin-coupled protein accumulation in the patient's tissues. In the patient's skin and B cells, IL-6 was highly expressed, and there was reduced expression of PSMB8. Downregulation of PSMB8 inhibited the differentiation of murine and human adipocytes in vitro, and injection of siRNA against Psmb8 in mouse skin reduced adipocyte tissue volume. These findings identify PSMB8 as an essential component and regulator not only of inflammation, but also of adipocyte differentiation, and indicate that immunoproteasomes have pleiotropic functions in maintaining the homeostasis of a variety of cell types.
Figures
References
-
- Dinarello CA. Interleukin-1beta and the autoinflammatory diseases. N Engl J Med. 2009;360(23):2467–2470. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
